
Η κατασκευή των Οργανισμών. Βιοχημική συνέχεια. Μέρος Τρίτο. Η ΧΗΜΙΚΗ ΚΑΙ ΒΙΟΧΗΜΙΚΗ ΕΞΕΛΙΞΗ ΤΟΥ ΚΥΤΤΑΡΟΥ: Ποια είναι και πως δημιουργούνται τα βασικά βιομορία των κυττάρων
Δήμητρα Σπανού, χημικός, καθηγήτρια στο 1ο Γυμνάσιο Δάφνης
ΤΑ ΚΥΡΙΑ ΒΙΟΛΟΓΙΚΑ ΜΟΡΙΑ ΤΩΝ ΟΡΓΑΝΙΣΜΩΝ
Πριν ακόμα εμφανιστεί η ζωή στον αρχέγονο πλανήτη Γη, υπήρχαν απλές χημικές ουσίες, όπως το υδρογόνο, το άζωτο, το μεθάνιο, η αμμωνία, το διοξείδιο του άνθρακα.
Σε ακραίες συνθήκες που επικρατούσαν τότε, πολλά από αυτά τα απλά μόρια συνδυάστηκαν και σχημάτισαν πολυπλοκότερα, όπως οι ενώσεις του άνθρακα, υδατάνθρακες, λιπαρά οξέα, ορισμένα αμινοξέα και βάσεις νουκλεϊκών οξέων.
Αυτά είναι πρόδρομα μόρια αποτέλεσαν τις βασικές ουσίες που εξελίχθηκαν σε μεγαλύτερες οργανικές ενώσεις που διακρίνονται από σταθερότητα, ποικιλομορφία και συμμετοχή στις λειτουργίες του κυττάρου, μπορούν γενικά να εντχθούν σε τέσσερες κατηγορίες:
Πρωτεινες, Λιπαρά , Υδατάνθρακες, Νουκλεικα οξέα και βέβαια το Νερό
Συγκρατούνται και αναπτύσσουν δυνάμεις μεταξύ τους μέσω Χημικών Δεσμών, οι οποίοι είναι :
Ιοντικός δεσμός, ομοιοπολικός δεσμός, δεσμός υδρογόνου, μη ομοιοπολικά δεσμοί wan der walls
Στην λειτουργία επίσης των βιολογικών μορίων συμμετέχουν και λειτουργικές ομάδες όπως:
Οι υδροξυλικές – ΟΗ (χαρακτηρίζονται από μεγάλη πολικότητα), καβονύλιο - C=O (συστατικό αλδεϋδών, κετονών ,δομικά συστατικά υδατανθράκων και λιπιδίων) καρβοξυλικές -COOH (μέρος των αμινοξέων και των οργανικών οξέων (ενεργειακά αποθέματα, ενδιάμεσα παράγωγα), αμινικές -NH2 (μέρος των αμινοξέων, ομοιοπολικοί δεσμοί) , φωσφορικές -PO4 (χημικές γέφυρες, ενεργοποίηση), σουλφυδρυλικές -SH(σταθεροποίηση)
ΤΙ ΕΙΝΑΙ ΟΙ ΠΡΩΤΕΪΝΕΣ ΤΟΥ ΚΥΤΤΑΡΟΥ
ΠΟΥ ΓΙΝΕΤΑΙ Η ΠΑΡΑΣΚΕΥΗ ΚΑΙ ΤΕΛΕΙΟΠΟΙΗΣΗ ΤΟΥΣ
Οι πρωτεϊνες είναι μεγάλα μόρια που αποτελούνται κατά κύριο λόγο από αμινοξέα. Ο αριθμός των αμινοξέων αυτών για μια πρωτεϊνη μπορεί να είναι από 3 έως 50000. Τα αμινοξέα που έχουν ανιχνευθεί είναι περίπου 170, όμως μόνο 20 από αυτά χρησιμοποιούνται για την κατασκευή πρωτεϊνών που ενώνονται μεταξύ τους σε διαφορετικούς συνδυασμούς, για ένα μεγάλο αριθμό πρωτεϊνών.
Ανάλογα με την διαμόρφωση των πρωτεϊνών στον χώρο διακρίνουμε τέσσερες δομές:
την Πρωτοταγή δομή των πρωτεϊνών που αντιπροσωπεύει την σειρά των αμινοξέων όπως τοποθετήθηκαν στα ριβοσώματα βάσει του γενετικού κώδικα συνδεδεμένα με ομοιοπολικούς δεσμούς,
την Δευτεροταγή δομή όπως διαμορφώηκε αυτή η σειρά στην συνέχεια με αναδιπλώσεις της αλυσίδας των αμινοξέων και σχηματισμό α έλικα, β νήμα με δεσμούς υδρογόνου,
την Τριτοταγή τρισδιάστατη διαμόρφωση σε μια πολυπλοκότερη διάταξη με δεσμούς υδρογόνου, δυνάμεις wan der walls κ.λ.π.
και την Τεταρτοταγή δομή με Πληρης τρισδιάστατη , διαμόρφωση συμπλόκου από δύο ή περισσότερες πολυπεπτιδικών αλύσίδων με δεσμούς υδρογόνου, δυνάμεις wan der walls κ.λ.π.
Η παρασκευή πρωτεϊνών στα κύτταρα αρχίζει από τα ριβοσωματα, που υπάρχουν είτε ελευθερα στο κυτταρόπλασμα ή προσκολλημένα στο αδρο ενδοπλασματικο δίκτυο στα ευκαρυωτικά κύτταρα
Στο προκαρυωτικα κυτταρο που διαθέτει ελεύθερα ριβοσώματα, σλλα όχι ενδοπλασματικό Δίκτυο, Golgi, Συσκευη Golgi, η σύνθεση πρωτεινων περιοριζεται στα ριβοσώματα.
Στο ευκαρυωτικό κύτταρο, η επεξεργασία και η τροποποίηση των πρωτεϊνών τους ολοκληρωνεται στο αδρό ενδοπλασματικό δίκτυ και στο Συστημα Golgi, στο οποίο τις προωθεί το ενδοπλασματικό δίκτυο. (Τα ριβοσώματα συνδέονται με πρωτεινες- υποδοχείς στην επιφάνεια του αδρου ενδοπλασματικου δικτύου ( ριβοφορίνες) . Η συνθεση των πρωτεϊνών συνεχίζεται έτσι μεσα στο αδρο ενδοπλασματικό δίκτυο).
Το αδρό ενδοπλασματικό δίκτυο αναγνωριζει, ταξινομεί και προωθεί τις διαφορετικές πρωτεινες στον προορισμό τους, για να τελειοποιηθουν ή να βρουν τη θέση τους.
Την παραπερα εξέλιξη της πρωτεΐνης θα την οδηγήσει σε επόμενη θέση,
με την μεταφορά της στο σύστημα Golgi ή στα λυσσοσώματα ή την παραμονή τους στο ενδοπλασματικό δίκτυο, καθορίζει η παραπέρα προσθήκη μιας σηματοδοτικής αλληλουχίας που θεωρείται σαν η "ετικετα" της και ειναι η αλληλουχια των αμινοξέων τους στο Ν τελικό ακρο της, όπως συντίθεται στο ριβόσωμα.
Αλλά πριν ακομα αποσταλούν στον προορισμό τους κι ενω ακομα οι πρωτεινες βρίσκονται στο Αδρο Ενδοπλασματικό Δίκτυο υφίστανται ηδη μετασχηματισμούς φυσικούς και βιοχημικούς, όπως δίπλωση, σχηματισμός υπομονάδων, δισουλφιδικών δεσμών, γλυκοζυλίωση, προσθήκη γλυκολιπιδιων.
Πρωτεΐνες που προορίζονται για το Συστημα Golgi που πιθανόν να εξελιχθουν σε λιποπρωτεϊνες ή πρωτεινογλυκάνες της μεμβράνης, πρωτεϊνες των λυσοσωματων με ενζυμική δραση, εκκριτικές πρωτεινες που θα πακεταριστούν σε κυστίδια και
θα οδηγηθουν έξω από το κυτταρο ή και πρωτεινες του δικτύου αυτου.
Οσες πρωτεϊνες προορίζονται για τα λυσσοσωματα, τροποποιούνται με την προσάρτιση του σακχάρου 6 φωσφορική μαννόζη.
Τροποποίηση πρωτεϊνών γινεται επίσης και στο Συστημα Golgi κυριων για τια γλυκοπρωτεΐνες.
Πρωτεΐνες που πρόκειται να ενσωματωθούν στην κυτταρική μεμβρανη, συνήθως υφίστανται γλυκοζυλιωση δηλαδη προσθηκη υδρογονανθρακα στην πρωτεινη.
Ακομα υπαρχει ένα είδος τροποποίησης , η πρωτεόλυση, που με αυτήν η πρωτείνη κόβεται στα δύο και έτσι ενεργοποιείται .
Άλλα είδη τροποποίησης των πρωτεϊνών είναι η φωσφορυλιωση που ενεργοποιεί γενικά το κυτταρο, η μεθυλίωση κυριως των ιστονων /( γυρω τους τυλιγεται υο DNA) , που ενεργοποιεί και απενεργοποιει γονίδια.
ΠΟΥ ΓΙΝΕΤΑΙ Η ΠΑΡΑΣΚΕΥΗ ΤΩΝ ΛΙΠΙΔΙΩΝ ΤΟΥ ΚΥΤΤΑΡΟΥ
Για τα ευκαρυωτικά κύτταρα, η σύνθεση των λιπιδίων και ιδιαίτερα των φωσφολιπιδίων της μεμβράνης γίνεται στο Λειο Ενδοπλασματικό Δίκτυο που αποτελει συνέχεια του Αδρού χωρίς να διαθέτει ριβοσώματα
Το ΛΕΔ είναι ζωτικής σημασίας για το κύτταρο, καθώς είναι η κυτταρική περιοχή στην οποία όχι μόνο γίνεται η επεξεργασία των παραγόμενων πρωτεϊνών, αλλά επίσης και σύνθεση των λιπιδίων, φωσφολιπιδίων, σφιγγολιπιδίων, στεροειδών και ποσταγλανδίνων.

Έτσι το ΛΕΔ είναι άφθονο σε κύτταρα που κατέχουν ιδιαίτερο ρόλο στον μεταβολισμό - σύνθεση λιπιδίων, όπως τα κύτταρα των όρχεων και των ωοθηκών τα οποία είναι κύτταρα που παράγουν στεροειδή. Το ίδιο συμβαίνει και στα ηπατικά κύτταρα όπου τα αυξημένο Λείο Ενδοπλασματικό τους Δίκτυο, συμμετέχει στην σύνθεση λιποπρωτεϊνών.
Το ΛΕΔ που αποτελεί τμήμα του ενδοκυττάριου μεμβρανικού συστήματος και επικοινωνεί με το ΑΕΔ και τη συσκευή Golgi . Το λείο ΕΔ στην πλειονότητα των κυττάρων καταλαμβάνει σχετικά μικρή έκταση, (εξαίρεση τα κύτταρα που εκκρίνουν ή επεξεργάζονται λιπίδια). Ειδικότερα αποτελεί τη θέση της σύνθεσης των μεμβρανικών λιπιδίων και της επεξεργασίας των πρωτεϊνών.
Το λείο ενδοπλασματικό δίκτυο (εικ. 13) είναι ένα σύστημα ακανόνιστου σχήματος μεμβρανικών σωληναρίων και κυστιδίων που δεν είναι συνδεδεμένα με ριβοσώματα εφόσον στις μεμβράνες του δεν υπάρχουν οι ριβοφορίνες. Το ΛΕΔ είναι ζωτικής σημασίας για το κύτταρο, καθώς είναι η κυτταρική περιοχή στην οποία όχι μόνο γίνεται η επεξεργασία των παραγόμενων πρωτεϊνών, αλλά και η σύνθεση των λιπιδίων και ιδιαίτερα των φωσφολιπιδίων της μεμβράνης. Τα ένζυμα που είναι υπεύθυνα για τη σύνθεση των λιπιδίων εντοπίζονται στην εξωτερική (προς το κυτοσόλιο) επιφάνεια του λείου ΕΔ για να έχουν άμεση προσπέλαση στα πρόδρομα μόρια των λιπιδίων.
Μετά τη σύνθεση και την ενσωμάτωσή τους στη λιπιδική διπλοστιβάδα του έξω μεμβρανικού τμήματος του λείου ΕΔ, τα φωσφολιπίδια μεταπηδούν στο έσω μεμβρανικό τμήμα με τη βοήθεια ειδικών πρωτεϊνών μεταφοράς που ονομάζονται φλιπάσες (flipases).
Εξειδικευμένες λειτουργίες του ΛΕΔ που συμμετέχουν λιπίδια αποτελούν:
- Η οξείδωση, σύζευξη και μεθυλίωση , αντιδράσεις που χρησιμοποιούνται από τα ηπατικά κύτταρα για να εξουδετερώσουν λιποδιαλυτές τοξικές ενώσεις την αλκοόλη, διαφόρους τύπους εντομοκτόνων κλπ και ορμόνες.
- Με την επεξεργασία αυτή, κυρίως υδροξυλίωση κάνει τις ενώσεις αυτές πιο ευδιάλυτες στο νερό, για την ευκολότερη απομάκρυνσή τους
ΠΟΥ ΓΙΝΕΤΑΙ Η ΠΑΡΑΣΚΕΥΗ ΑΛΛΩΝ ΧΡΗΣΙΜΩΝ ΜΟΡΙΩΝ ΤΟΥ ΚΥΤΤΑΡΟΥ
Η παρασκευή των στεροειδών ορμονών του κυττάρου των σπονδυλωτών γίνεται επίσης στο λείο ενδοπλασματικό δίκτυο, ιδιαίτερα στο λείο Ενδοπλασματικό των ηπατικών κυττάρων.
Στην κατασκευή υδατανθράκων (που εμπλέκονται σε δομικές και λειτουργικές διεργασίες των κυττάρων π.χ. γλυκολιπίδια, γλυκοπρωτεϊνες, γλυκοκάλυκας) σχετίζεται το λείο ενδοπλασματικό δίκτυο, κυρίως των ηπατοκυττάρων, εφόσον εκεί βρίσκονται τα περισσότερα ένζυμα διάσπασης των υδατανθράκων αρχικά του κολαγόνου.

Δήμητρα Σπανού
Οι
ΠΗΓΕΣ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΗΛΕΚΤΡΟΛΟΓΩΝ ΜΗΧΑΝΙΚΩΝ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΥΠΟΛΟΓΙΣΤΩΝ
ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ ΚΡΟΥΣΟΥΛΟΥΔΗ ΜΑΡΙΑ ΤΟΥ ΝΙΚΟΛΑΟΥ ΑΜ 22809
intracellular_communication
Ενδοπλασματικό δίκτυο - Βικιπαίδεια (wikipedia.org)
ΕΙΣΑΓΩΓΗ ΣΤΗΝ ΚΥΤΤΑΡΙΚΗ ΒΙΟΛΟΓΙΑ ΚΑΙ ΓΕΝΕΤΙΚΗ. Σαρρής Παναγιώτης Μοριακός Βιολόγος. Κλινική Χημεία MSc Μοριακή Μικροβιολογία- Βιοτεχνολογία MSc MOLBIOL_Sarris.pdf
https://studyflix.de/biologie/ribosomen-1894
ακατέργαστο
Όλα τα κύτταρα του σώματός μας είναι ίδια μεταξύ τους; - Mothersblog.gr
Το ΛΕΔ αποτελεί τμήμα του ενδοκυττάριου μεμβρανικού συστήματος και επικοινωνεί με το ΑΕΔ και τη συσκευή Golgi . Το λείο ΕΔ στην πλειονότητα των κυττάρων καταλαμβάνει σχετικά μικρή έκταση, με εξαίρεση τα κύτταρα που εκκρίνουν ή επεξεργάζονται λιπίδια. Ειδικότερα αποτελεί τη θέση της σύνθεσης των μεμβρανικών λιπιδίων και της επεξεργασίας των πρωτεϊνών.
Το λείο ενδοπλασματικό δίκτυο (εικ. 13) είναι ένα σύστημα ακανόνιστου σχήματος μεμβρανικών σωληναρίων και κυστιδίων που δεν είναι συνδεδεμένα με ριβοσώματα. Το ΛΕΔ είναι ζωτικής σημασίας για το κύτταρο, καθώς είναι η κυτταρική περιοχή στην οποία όχι μόνο γίνεται η επεξεργασία των παραγόμενων πρωτεϊνών, αλλά και η σύνθεση των λιπιδίων και ιδιαίτερα των φωσφολιπιδίων της μεμβράνης. Τα ένζυμα που είναι υπεύθυνα για τη σύνθεση των λιπιδίων εντοπίζονται στην εξωτερική (προς το κυτοσόλιο) επιφάνεια του λείου ΕΔ για να έχουν άμεση προσπέλαση στα πρόδρομα μόρια των λιπιδίων.
Μετά τη σύνθεση και την ενσωμάτωσή τους στη λιπιδική διπλοστιβάδα του έξω μεμβρανικού τμήματος του λείου ΕΔ, τα φωσφολιπίδια μεταπηδούν στο έσω μεμβρανικό τμήμα με τη βοήθεια ειδικών πρωτεϊνών μεταφοράς που ονομάζονται φλιπάσες (flipases).
Εξειδικευμένες λειτουργίες του ΑΕΔ αποτελούν:
- Η διάσπαση του γλυκογόνου στα ηπατικά κύτταρα
- Η οξείδωση, σύζευξη και μεθυλίωση , αντιδράσεις που χρησιμοποιούνται από τα ηπατικά κύτταρα για να εξουδετερώσουν ορμόνες, την αλκοόλη, διαφόρους τύπους εντομοκτόνων κλπ
Η συμμετοχή στο μηχανισμό συστολής των μυϊκών κυττάρων, όπου αναγνωρίζεται με μια εξειδικευμένη μορφή που ονομάζεται σαρκοπλασματικό δίκτυο και συμμετέχει στη δέσμευση και απελευθέρωση των ιόντων ασβεστίου, τα οποία ρυθμίζουν τη μυϊκή συστολή.
Οι δύο μορφές διαφέρουν επίσης ως προς τη λειτουργία τους. Το ακατέργαστο ER εμπλέκεται κυρίως στην παραγωγή (βιοσύνθεση πρωτεϊνών) και την αναδίπλωση των πρωτεϊνών. Το καθήκον του λείου ER είναι να παράγει διάφορες ουσίες όπως λιπαρά οξέα ή ορμόνες και να αποτοξινώνει το κύτταρο.
https://studyflix.de/biologie/ribosomen-1894
Η προηγούμενη ενότητα ανασκόπησε τις κύριες μεταβολικές αντιδράσεις με τις οποίες το κύτταρο αποκτά και αποθηκεύει ενέργεια με τη μορφή ATP. Αυτή η μεταβολική ενέργεια χρησιμοποιείται στη συνέχεια για την ολοκλήρωση διαφόρων εργασιών, συμπεριλαμβανομένης της σύνθεσης μακρομορίων και άλλων κυτταρικών συστατικών. Έτσι, η ενέργεια που προέρχεται από τη διάσπαση των οργανικών μορίων (καταβολισμός) χρησιμοποιείται για να οδηγήσει τη σύνθεση άλλων απαιτούμενων συστατικών του κυττάρου. Τα περισσότερα καταβολικά μονοπάτια περιλαμβάνουν την οξείδωση οργανικών μορίων που συνδέονται με την παραγωγή ενέργειας (ATP) και αναγωγικής ισχύος (NADH). Αντίθετα, οι βιοσυνθετικές (αναβολικές) οδοί γενικά περιλαμβάνουν τη χρήση τόσο του ATP όσο και της αναγωγικής ισχύος (συνήθως με τη μορφή NADPH) για την παραγωγή νέων οργανικών ενώσεων. Μια κύρια βιοσυνθετική οδός, η σύνθεση υδατανθράκων από CO 2 και H 2Το O κατά τις σκοτεινές αντιδράσεις της φωτοσύνθεσης , συζητήθηκε στην προηγούμενη ενότητα. Πρόσθετες οδοί που οδηγούν στη βιοσύνθεση των κύριων κυτταρικών συστατικών (υδατάνθρακες, λιπίδια , πρωτεΐνες και νουκλεϊκά οξέα) εξετάζονται στις ενότητες που ακολουθούν.
Εκτός από το ότι λαμβάνεται απευθείας από τα τρόφιμα ή παράγεται με φωτοσύνθεση , η γλυκόζη μπορεί να συντεθεί και από άλλα οργανικά μόρια. Στα ζωικά κύτταρα, η σύνθεση γλυκόζης ( γλυκονεογένεση ) συνήθως ξεκινά με γαλακτικό (που παράγεται με αναερόβια γλυκόλυση ), αμινοξέα (που προέρχονται από τη διάσπαση πρωτεϊνών ) ή γλυκερίνη (που παράγεται από τη διάσπαση των λιπιδίων ). Τα φυτά (αλλά όχι τα ζώα) μπορούν επίσης να συνθέσουν γλυκόζη από λιπαρά οξέα —μια διαδικασία που είναι ιδιαίτερα σημαντική κατά τη βλάστηση των σπόρων, όταν η ενέργεια αποθηκεύεται ως λίπηπρέπει να μετατραπεί σε υδατάνθρακες για να υποστηρίξει την ανάπτυξη του φυτού. Τόσο στα ζωικά όσο και στα φυτικά κύτταρα, τα απλά σάκχαρα πολυμερίζονται και αποθηκεύονται ως πολυσακχαρίτες.
Η γλυκονεογένεση περιλαμβάνει τη μετατροπή του πυροσταφυλικού σε γλυκόζη - ουσιαστικά το αντίστροφο της γλυκόλυσης . Ωστόσο, όπως συζητήθηκε προηγουμένως, η γλυκολυτική μετατροπή της γλυκόζης σε πυροσταφυλικό είναι μια οδός παραγωγής ενέργειας, δημιουργώντας δύο μόρια από το καθένα ATP και NADH. Αν και ορισμένες αντιδράσεις γλυκόλυσης είναι άμεσα αναστρέψιμες, άλλες θα προχωρήσουν μόνο προς την κατεύθυνση της διάσπασης της γλυκόζης, επειδή σχετίζονται με μεγάλη μείωση της ελεύθερης ενέργειας. Αυτές οι ενεργειακά ευνοϊκές αντιδράσεις γλυκόλυσης παρακάμπτονται κατά τη διάρκεια της γλυκονεογένεσης από άλλες αντιδράσεις (που καταλύονται από διαφορετικά ένζυμα) που συνδέονται με τη δαπάνη του ATP και του NADH προκειμένου να τα οδηγήσουν προς την κατεύθυνση της σύνθεσης γλυκόζης. Συνολικά, η παραγωγή γλυκόζης από δύο μόρια πυροσταφυλικού άλατος απαιτεί τέσσερα μόρια ATP, δύο GTP και δύο NADH. Αυτή η διαδικασία είναι πολύ πιο δαπανηρή από την απλή αντιστροφή της γλυκόλυσης (η οποία θα απαιτούσε δύο μόρια ATP και δύο NADH), απεικονίζοντας την πρόσθετη ενέργεια που απαιτείται για να οδηγήσει το μονοπάτι προς την κατεύθυνση της βιοσύνθεσης.
Τόσο στα φυτικά όσο και στα ζωικά κύτταρα, η γλυκόζη αποθηκεύεται με τη μορφή πολυσακχαριτών ( άμυλο και γλυκογόνο , αντίστοιχα). Η σύνθεση πολυσακχαριτών, όπως και όλων των άλλων μακρομορίων, είναι μια αντίδραση που απαιτεί ενέργεια. Όπως σημειώθηκε προηγουμένως, η σύνδεση δύο σακχάρων με έναν γλυκοσιδικό δεσμό μπορεί να γραφτεί ως αντίδραση αφυδάτωσης, στην οποία αφαιρείται το H2O (βλ. Εικόνα 2.3 ). Μια τέτοια αντίδραση, ωστόσο, είναι ενεργειακά δυσμενής και ως εκ τούτου δεν μπορεί να προχωρήσει προς τα εμπρός. Κατά συνέπεια, ο σχηματισμός ενός γλυκοσιδικού δεσμού πρέπει να συζευχθεί με μια αντίδραση παραγωγής ενέργειας, η οποία επιτυγχάνεται με τη χρήση νουκλεοτιδικών σακχάρων ως ενδιάμεσων σε πολυσακχαρίτεςσύνθεση ( Εικόνα 2.40 ). Η γλυκόζη αρχικά φωσφορυλιώνεται σε μια αντίδραση που καθοδηγείται από το ATP στη 6-φωσφορική γλυκόζη, η οποία στη συνέχεια μετατρέπεται σε 1-φωσφορική γλυκόζη. Η γλυκόζη-1-φωσφορική αντιδρά με την UTP (τριφωσφορική ουριδίνη), δίνοντας UDP-γλυκόζη συν πυροφωσφορικό, το οποίο υδρολύεται σε φωσφορικό άλας με την απελευθέρωση πρόσθετης ελεύθερης ενέργειας. Η UDP-γλυκόζη είναι ένα ενεργοποιημένο ενδιάμεσο που στη συνέχεια δίνει το υπόλειμμα γλυκόζης του σε μια αναπτυσσόμενη αλυσίδα πολυσακχαρίτη σε μια ενεργειακά ευνοϊκή αντίδραση. Έτσι, η χημική ενέργεια με τη μορφή ATP και UTP οδηγεί τη σύνθεση πολυσακχαριτών από απλά σάκχαρα.

Εικόνα 2.40
Σύνθεση πολυσακχαριτών. Η γλυκόζη μετατρέπεται αρχικά σε μια ενεργοποιημένη μορφή, την UDP-γλυκόζη, με κόστος ενός μορίου το καθένα από τα ATP και UTP. Το υπόλειμμα γλυκόζης μπορεί στη συνέχεια να μεταφερθεί από την UDP-γλυκόζη σε μια αναπτυσσόμενη αλυσίδα πολυσακχαρίτη σε μια ενεργειακή (περισσότερα...)
Λιπίδια
Τα λιπίδια είναι σημαντικά μόρια αποθήκευσης ενέργειας και το κύριο συστατικό των κυτταρικών μεμβρανών. Συντίθενται από το ακετύλιο CoA, το οποίο σχηματίζεται από τη διάσπαση των υδατανθράκων, σε μια σειρά αντιδράσεων που μοιάζουν με το αντίστροφο της οξείδωσης των λιπαρών οξέων. Όπως και με τη βιοσύνθεση υδατανθράκων , ωστόσο, οι αντιδράσεις που οδηγούν στη σύνθεση λιπαρών οξέωνδιαφέρουν από εκείνα που εμπλέκονται στην αποδόμησή τους και οδηγούνται προς τη βιοσυνθετική κατεύθυνση, καθώς συνδέονται με τη δαπάνη τόσο της ενέργειας με τη μορφή ATP όσο και με τη μείωση της ισχύος με τη μορφή NADPH. Τα λιπαρά οξέα συντίθενται με τη σταδιακή προσθήκη μονάδων δύο άνθρακα που προέρχονται από το ακετύλιο CoA σε μια αναπτυσσόμενη αλυσίδα. Η προσθήκη καθεμιάς από αυτές τις μονάδες δύο άνθρακα απαιτεί τη δαπάνη ενός μορίου ATP και δύο μορίων NADPH.
Το κύριο προϊόν της βιοσύνθεσης λιπαρών οξέων, που εμφανίζεται στο κυτταρόπλασμα των ευκαρυωτικών κυττάρων , είναι το παλμιτικό λιπαρό οξύ 16 άνθρακα. Τα κύρια συστατικά των κυτταρικών μεμβρανών ( φωσφολιπίδια , σφιγγομυελίνη και γλυκολιπίδια) συντίθενται στη συνέχεια από ελεύθερα λιπαρά οξέα στο ενδοπλασματικό δίκτυο και στη συσκευή Golgi (βλ. Κεφάλαιο 9).
Σε αντίθεση με τους υδατάνθρακες και τα λιπίδια , οι πρωτεΐνες (καθώς και τα νουκλεϊκά οξέα) περιέχουν άζωτο εκτός από άνθρακα, υδρογόνο και οξυγόνο. Το άζωτο ενσωματώνεται σε οργανικές ενώσεις από διαφορετικές πηγές σε διαφορετικούς οργανισμούς ( Εικόνα 2.41 ). Ορισμένα βακτήρια μπορούν να χρησιμοποιήσουν το ατμοσφαιρικό N 2 με μια διαδικασία που ονομάζεται δέσμευση αζώτου , κατά την οποία το N 2 μειώνεται σε NH 3 σε βάρος της ενέργειας με τη μορφή ATP. Αν και σχετικά λίγα είδη βακτηρίων είναι ικανά για δέσμευση αζώτου , τα περισσότερα βακτήρια, μύκητες και φυτά μπορούν να χρησιμοποιήσουν νιτρικό άλας (NO 3 - ), το οποίο είναι ένα κοινό συστατικό του εδάφους, μειώνοντάς το σε NH3 μέσω ηλεκτρονίων που προέρχονται από NADH ή NADPH. Τέλος, όλοι οι οργανισμοί είναι σε θέση να ενσωματώνουν αμμωνία (NH 3 ) σε οργανικές ενώσεις.

Εικόνα 2.41
Αφομοίωση του αζώτου σε οργανικές ενώσεις. Η αμμωνία ενσωματώνεται στις οργανικές ενώσεις από όλους τους οργανισμούς. Ορισμένα βακτήρια είναι ικανά να μετατρέψουν το ατμοσφαιρικό άζωτο σε αμμωνία και τα περισσότερα βακτήρια, μύκητες και φυτά μπορούν να χρησιμοποιήσουν νιτρικά άλατα από το έδαφος. (περισσότερο...)
Το NH 3 ενσωματώνεται σε οργανικά μόρια κυρίως κατά τη σύνθεση των αμινοξέων γλουταμικό και γλουταμίνη, τα οποία προέρχονται από το ενδιάμεσο α-κετογλουταρικό του κύκλου του κιτρικού οξέος . Αυτά τα αμινοξέα στη συνέχεια χρησιμεύουν ως δότες αμινομάδων κατά τη διάρκεια της σύνθεσης των άλλων αμινοξέων, τα οποία επίσης προέρχονται από κεντρικές μεταβολικές οδούς, όπως η γλυκόλυση και ο κύκλος του κιτρικού οξέος ( Εικόνα 2.42 ). Η πρώτη ύλη για το αμινοξύΗ σύνθεση λαμβάνεται έτσι από τη γλυκόζη και τα αμινοξέα συντίθενται με κόστος τόσο της ενέργειας (ATP) όσο και της αναγωγικής ισχύος (NADPH). Πολλά βακτήρια και φυτά μπορούν να συνθέσουν και τα 20 αμινοξέα. Οι άνθρωποι και άλλα θηλαστικά, ωστόσο, μπορούν να συνθέσουν μόνο περίπου τα μισά από τα απαιτούμενα αμινοξέα. Το υπόλοιπο πρέπει να λαμβάνεται από διαιτητικές πηγές ( Πίνακας 2.2 ).
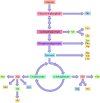
Εικόνα 2.42
Βιοσύνθεση αμινοξέων. Οι σκελετοί άνθρακα των αμινοξέων προέρχονται από ενδιάμεσα στη γλυκόλυση και στον κύκλο του κιτρικού οξέος.
https://www.ncbi.nlm.nih.gov/books/NBK9956/
Εικόνα 2.40
Σύνθεση πολυσακχαριτών. Η γλυκόζη μετατρέπεται αρχικά σε μια ενεργοποιημένη μορφή, την UDP-γλυκόζη, με κόστος ενός μορίου το καθένα από τα ATP και UTP. Το υπόλειμμα γλυκόζης μπορεί στη συνέχεια να μεταφερθεί από την UDP-γλυκόζη σε μια αναπτυσσόμενη αλυσίδα πολυσακχαρίτη σε μια ενεργειακή (περισσότερα...)
Λιπίδια
Τα λιπίδια είναι σημαντικά μόρια αποθήκευσης ενέργειας και το κύριο συστατικό των κυτταρικών μεμβρανών. Συντίθενται από το ακετύλιο CoA, το οποίο σχηματίζεται από τη διάσπαση των υδατανθράκων, σε μια σειρά αντιδράσεων που μοιάζουν με το αντίστροφο της οξείδωσης των λιπαρών οξέων. Όπως και με τη βιοσύνθεση υδατανθράκων , ωστόσο, οι αντιδράσεις που οδηγούν στη σύνθεση λιπαρών οξέωνδιαφέρουν από εκείνα που εμπλέκονται στην αποδόμησή τους και οδηγούνται προς τη βιοσυνθετική κατεύθυνση, καθώς συνδέονται με τη δαπάνη τόσο της ενέργειας με τη μορφή ATP όσο και με τη μείωση της ισχύος με τη μορφή NADPH. Τα λιπαρά οξέα συντίθενται με τη σταδιακή προσθήκη μονάδων δύο άνθρακα που προέρχονται από το ακετύλιο CoA σε μια αναπτυσσόμενη αλυσίδα. Η προσθήκη καθεμιάς από αυτές τις μονάδες δύο άνθρακα απαιτεί τη δαπάνη ενός μορίου ATP και δύο μορίων NADPH.
Το κύριο προϊόν της βιοσύνθεσης λιπαρών οξέων, που εμφανίζεται στο κυτταρόπλασμα των ευκαρυωτικών κυττάρων , είναι το παλμιτικό λιπαρό οξύ 16 άνθρακα. Τα κύρια συστατικά των κυτταρικών μεμβρανών ( φωσφολιπίδια , σφιγγομυελίνη και γλυκολιπίδια) συντίθενται στη συνέχεια από ελεύθερα λιπαρά οξέα στο ενδοπλασματικό δίκτυο και στη συσκευή Golgi (βλ. Κεφάλαιο 9).