
Χαρακτηριστικά της δομής των πλασματικών μεμβρανών σε διάφορους οργανισμούς
Χαρακτηριστικά της δομής των πλασματικών μεμβρανών σε διάφορους οργανισμούς.
Μεμβράνες φυτικών κυττάρων Δομή. Το πάχος των βιομεμβρανών δεν ξεπερνά τα 6-10 nm. Σύμφωνα με την υπόθεση του υγρού μωσαϊκού της δομής των βιολογικών μεμβρανών, η βάση της μεμβράνης είναι ένα διπλό στρώμα φωσφολιπιδίων με μια ορισμένη ποσότητα άλλων λιπιδίων (γαλακτολιπίδια, στερόλες, λιπαρά οξέα κ.λπ.) και τα λιπίδια στρέφονται προς το καθένα. άλλα με τα υδρόφοβα άκρα τους (Εικόνα 9). Περιοχές μορίων πολικών λιπιδίων, που πολύ συχνά σχηματίζονται από ακόρεστα λιπαρά οξέα, και στερόλες παρέχουν μια κάπως χαλαρή (υγρή) κατάσταση της διπλής στοιβάδας. Το πολικό τμήμα του μορίου των φωσφολιπιδίων αλληλεπιδρά καλά με το υδάτινο περιβάλλον. Ο πολλαπλός δεσμός στη μάλλον μακριά μη πολική «ουρά» των φωσφολιπιδίων βρίσκεται στη διαμόρφωση cis, επομένως το «πόδι» δεν είναι ίσιο, αλλά, όπως ήταν, κυρτό (Εικόνα 10 α).
Εάν ένα δείγμα ενός φωσφολιπιδίου αναδευτεί σε ένα υδατικό μέσο, σχηματίζονται τα λεγόμενα μικκύλια (Εικόνα 10 β) κατασκευασμένα με τέτοιο τρόπο ώστε οι πολικές κεφαλές να μετατρέπονται στο υδατικό στρώμα και οι μη πολικές ουρές να βρίσκονται μέσα στα μικκύλια .
Η ικανότητα ορισμένων λιπιδίων να «αυτοσυναρμολογούνται» σε διπλές στιβάδες είναι μια πολύ σημαντική ιδιότητα που παίζει καθοριστικό ρόλο στην κατασκευή των κυτταρικών μεμβρανών. που περιβάλλονται από μεμβράνες και διαφέρουν ως προς τον βαθμό δραστηριότητας των χημικών ενώσεων που περιέχονται σε αυτές και τα συστήματα που ρυθμίζουν τους μετασχηματισμούς τους. Τα διαμερίσματα ενός ευκαρυωτικού κυττάρου ονομάζονται οργανίδια. Μονομεμβρανικά δομικά συστατικά των κυττάρων: πλασμάλημμα, ενδοπλασματικό δίκτυο, σύμπλεγμα Golgi, λυσοσώματα, κενοτόπιο.
Δομικά συστατικά δύο μεμβρανών κυττάρων: πυρήνας, μιτοχόνδρια, πλαστίδια.
2) Λειτουργία φραγμού . Για τα κύτταρα και τα υποκυτταρικά σωματίδια, οι μεμβράνες χρησιμεύουν ως μηχανικό φράγμα που τα χωρίζει από τον εξωτερικό χώρο.
3) Έλεγχος απορρόφησης και έκκρισης ουσιών (λειτουργία μεταφοράς). Η πλασματική μεμβράνη περιέχει πολλά συστήματα μεταφοράς, κυρίως για τη μεταφορά ιόντων - διαύλων ιόντων, φορέων ιόντων και αντλιών ιόντων. Χάρη σε αυτά, πραγματοποιείται μια πολύ ακριβής και επιλεκτική μεταφορά ιόντων τόσο μέσα στο κύτταρο όσο και έξω, δηλ. βέλτιστη παροχή των κυττάρων με τα απαραίτητα ιόντα.
4) Συσσώρευση και μετατροπή ενέργειας. Το πλάσμα οποιουδήποτε κυττάρου είναι μια ενεργοποιημένη μεμβράνη, δηλαδή υπάρχει μια βαθμίδα ηλεκτροχημικού δυναμικού Δμ H+ , η οποία χρησιμοποιείται για την εκτέλεση χρήσιμης εργασίας, κυρίως για την ενεργό μεταφορά ουσιών μέσω της μεμβράνης.
5) Τοποθέτηση και διασφάλιση της εργασίας των ενζύμων. Η πλασματική μεμβράνη περιέχει πολλά ένζυμα. Αυτά είναι ένζυμα κατασκευής κυτταρικού τοιχώματος (συνθάση κυτταρίνης), ένζυμα συστημάτων σηματοδότησης (φωσφολιπάσες C, Α, αδενυλική κυκλάση)
6) Λειτουργία υποδοχέα. Το κυτταρικό πλάσμα είναι ένα μωσαϊκό από διάφορους υποδοχείς για ενδογενή σήματα (κυρίως φυτοορμόνες) και εξωτερικές επιδράσεις. Η θερμοκρασία αλλάζει την ακαμψία (ρευστότητα) της μεμβράνης, δηλ. τις μηχανικές του ιδιότητες. Μια αλλαγή σε αυτές τις ιδιότητες της πλασματικής μεμβράνης οδηγεί στο άνοιγμα ή το κλείσιμο των διαύλων μηχανοαισθητήρων ιόντων που βρίσκονται σε αυτήν.
7) Λειτουργία σήματος. Πολλά συστατικά του πλάσματος μετά την αντίληψη των σημάτων χρησιμεύουν ως πηγή δευτερευόντων αγγελιοφόρων - μορίων που «μεταδίδουν» το σήμα κατά μήκος της κούρσας αναμετάδοσης και το ενισχύουν.
Το κυτταρικό τοίχωμα παρέχει μηχανική αντοχή στο στοιχείο, δίνοντάς του μια άκαμπτη (άκαμπτη) δομή, έτσι ώστε το κύτταρο να μπορεί να αντέξει υψηλή εσωτερική οσμωτική πίεση (5-20 MPa). Επιπλέον, το κυτταρικό τοίχωμα μπορεί να παρέχει κάποιο βαθμό επιλεκτικής διαπερατότητας σε ουσίες μικρού μοριακού βάρους, καθώς και την ικανότητα να αλληλεπιδρά με άλλα κύτταρα, ιούς και φυσικές επιφάνειες. Η δομή του κυτταρικού τοιχώματος σε διαφορετικούς οργανισμούς έχει τα δικά της χαρακτηριστικά.
Τα κύτταρα των περισσότερων ιστών των πολυκύτταρων ζώων δεν περιέχουν έντονο κυτταρικό τοίχωμα. Τα φυτικά κύτταρα, αντίθετα, έχουν ένα πολύ περίπλοκο κυτταρικό τοίχωμα κατασκευασμένο από μικροϊνίδια κυτταρίνης ενσωματωμένα σε μια μήτρα (πηκτίνης και ημικυτταρινών).
Τα κυτταρικά τοιχώματα των ζυμομυκήτων και των νηματοειδών μυκήτων αποτελούνται από ομο- και ετεροπολυσακχαρίτες (γλυκάνες, χιτίνη) και ένα σύμπλεγμα πρωτεϊνών.Το πάχος αυτών των στρωμάτων φτάνει το 1 μm.
Το κυτταρικό τοίχωμα των θετικών στον κεραυνό βακτηρίων είναι κατασκευασμένο κυρίως από γέφυρες αμινοξέων που περιέχουν ετεροπολυσακχαρίτη μουρεΐνης. Στα gram-αρνητικά βακτήρια, το στρώμα μουρεΐνης είναι μικρό, αλλά το κυτταρικό τοίχωμα περιέχει μια εξωτερική μεμβράνη φτιαγμένη από φωσφολιπίδια, πρωτεΐνες και λιποπολυσακχαρίτη, η οποία παρέχει κάποιο βαθμό επιλεκτικής διαπερατότητας και περιέχει υποδοχείς φάγου και αντιγόνα. Το πάχος του κυτταρικού τοιχώματος κυμαίνεται από 15 έως 80 nm
Δομές ενεργές με μεμβράνες.
Επί του παρόντος, έχει αποδειχθεί η ύπαρξη τριών τύπων τέτοιων πρωτεϊνών μεταφοράς: κανάλια, φορείς και αντλίες.
Каналы - это трансмембранные белки, которые действуют как поры. Иногда их называют селективными фильтрами. Μεταφορά через каналы, как правило, пассивный. Специфичность транспортируемого вещества определяется свойствами поверхности поры. Как правило, через каналы передвигаются ионы. Скорость транспорта εξαρτάται од их величины и заряда. Если пора открыта, то вещества проходят быстро. Однако каналы открыты не всегда. Имеется механизм «ворот», который под влиянием внешнего сигнала открывает или закрывает канал. Долгое время представлялась труднообъяснимой высокая проницаемость мембраны (10 mkm/s) για воды - πλεονεκτήματα полярного и нерастворимого в липидах. В настоящее время открыты ενσωματωμένες μεμβράνες, προμηθευτές κανάλι через μεμβράνη для проникновения воды - ακουαρίνα. Η ικανότητα των ακουαπορινών να μεταφέρουν νερό ρυθμίζεται από τη διαδικασία της φωσφορυλίωσης. Η προσκόλληση και η δωρεά φωσφορικών ομάδων σε ορισμένα αμινοξέα ακουαπορίνης έχει αποδειχθεί ότι επιταχύνει ή αναστέλλει την είσοδο νερού, αλλά δεν επηρεάζει την κατεύθυνση μεταφοράς.
Οι φορείς είναι συγκεκριμένες πρωτεΐνες που μπορούν να συνδεθούν με μια φερόμενη ουσία. Στη δομή αυτών των πρωτεϊνών υπάρχουν ομαδοποιήσεις που είναι προσανατολισμένες με συγκεκριμένο τρόπο προς την εξωτερική ή την εσωτερική επιφάνεια. Ως αποτέλεσμα μιας αλλαγής στη διαμόρφωση των πρωτεϊνών, η ουσία μεταφέρεται προς τα έξω ή προς τα μέσα. Δεδομένου ότι για τη μεταφορά κάθε μεμονωμένου μορίου ή ιόντος, ο φορέας πρέπει να αλλάξει τη διάταξή του, ο ρυθμός μεταφοράς μιας ουσίας είναι αρκετές φορές χαμηλότερος από τη μεταφορά μέσω καναλιών. Η παρουσία πρωτεϊνών μεταφοράς φάνηκε όχι μόνο στο πλάσμα, αλλά και στον τονοπλάστη. Η μεταφορά του μεταφορέα μπορεί να είναι ενεργή ή παθητική. Στην τελευταία περίπτωση, μια τέτοια μεταφορά πηγαίνει προς την κατεύθυνση του ηλεκτροχημικού δυναμικού και δεν απαιτεί ενέργεια. Αυτός ο τύπος μεταφοράς ονομάζεται διευκολυνόμενη διάχυση. Χάρη στους μεταφορείς, πηγαίνει με μεγαλύτερη ταχύτητα,
Σύμφωνα με την έννοια του έργου των φορέων, το ιόν (Μ) αντιδρά με τον φορέα του (Χ) στην επιφάνεια της μεμβράνης ή κοντά σε αυτήν. Αυτή η πρώτη αντίδραση μπορεί να περιλαμβάνει είτε προσρόφηση ανταλλαγής είτε κάποιο είδος χημικής αλληλεπίδρασης. Ούτε ο ίδιος ο φορέας ούτε το σύμπλοκό του με το ιόν μπορούν να περάσουν στο εξωτερικό περιβάλλον. Ωστόσο, το σύμπλεγμα μεταφορέα ιόντων (MX) είναι κινητό εντός της ίδιας της μεμβράνης και κινείται προς την αντίθετη πλευρά της. Εδώ, αυτό το σύμπλοκο αποσυντίθεται και απελευθερώνει ένα ιόν στο εσωτερικό περιβάλλον για να σχηματίσει έναν πρόδρομο φορέα (X´). Αυτός ο πρόδρομος φορέας ταξιδεύει και πάλι προς το εξωτερικό της μεμβράνης, όπου μετατρέπεται και πάλι από πρόδρομο σε φορέα που μπορεί να συνδυαστεί με άλλο ιόν στην επιφάνεια της μεμβράνης. Όταν μια ουσία ικανή να σχηματίσει ένα σταθερό σύμπλοκο με έναν φορέα εισάγεται στο μέσο, η μεταφορά της ουσίας εμποδίζεται. Πειράματα που έγιναν σε τεχνητές μεμβράνες λιπιδίων έδειξαν ότι η μεταφορά ιόντων μπορεί να πραγματοποιηθεί υπό την επίδραση ορισμένων αντιβιοτικών που παράγονται από βακτήρια και μύκητες - ιονοφόρα (Εικόνα 12). Η μεταφορά με τη συμμετοχή φορέων έχει την ιδιότητα του κορεσμού, δηλαδή, με την αύξηση της συγκέντρωσης των ουσιών στο περιβάλλον διάλυμα, ο ρυθμός εισόδου πρώτα αυξάνεται και μετά παραμένει σταθερός. Αυτό οφείλεται στον περιορισμένο αριθμό των μεταφορέων. Οι φορείς είναι συγκεκριμένοι, δηλαδή εμπλέκονται στη μεταφορά ορισμένων μόνο ουσιών και, ως εκ τούτου, διασφαλίζουν την επιλεκτικότητα της πρόσληψης. Αυτό δεν αποκλείει την πιθανότητα ο ίδιος φορέας να μπορεί να μεταφέρει πολλά ιόντα. Για παράδειγμα, ο μεταφορέας K + , ο οποίος είναι ειδικός για αυτό το ιόν, μεταφέρει επίσης Rb + και Na + , αλλά δεν μεταφέρει Cl- ή αφόρτιστα μόρια σακχαρόζης. Μια πρωτεΐνη μεταφοράς ειδική για ουδέτερα οξέα ανέχεται τα αμινοξέα γλυκίνη, βαλίνη, αλλά όχι ασπαραγίνη ή λυσίνη. Λόγω της ποικιλομορφίας και της ειδικότητας των πρωτεϊνών, πραγματοποιείται η εκλεκτική τους αντίδραση με ουσίες του περιβάλλοντος και, ως εκ τούτου, η επιλεκτική μεταφορά τους.
Οι αντλίες (αντλίες) είναι αναπόσπαστες πρωτεΐνες μεταφοράς που παρέχουν ενεργά ιόντα. Ο όρος «αντλία» υποδηλώνει ότι η ροή είναι με την κατανάλωση ελεύθερης ενέργειας και αντίθετα με την ηλεκτροχημική κλίση. Η ενέργεια που χρησιμοποιείται για την ενεργό είσοδο ιόντων
Τα λιπίδια που αποτελούν τη διπλή στιβάδα της μεμβράνης δεν είναι σταθερά σταθερά, αλλά αλλάζουν συνεχώς θέσεις. Οι κινήσεις των μορίων των λιπιδίων είναι δύο τύπων:
εντός της μονοστιβάδας του (πλευρική διάχυση).
με την αναδιάταξη δύο μορίων λιπιδίων που αντιτίθενται μεταξύ τους σε δύο μονοστοιβάδες («flip flop»).
Κατά τη διάρκεια της πλευρικής διάχυσης, τα μόρια λιπιδίων υφίστανται εκατομμύρια μεταθέσεις ανά δευτερόλεπτο και η ταχύτητά τους είναι 5-10 μm/s. Οι αναδιατάξεις των μορίων λιπιδίων από τη μία μονοστιβάδα στην άλλη συμβαίνουν πολύ λιγότερο συχνά, αλλά μπορούν να επιταχυνθούν από τις πρωτεΐνες της μεμβράνης.
Οι μεμβράνες περιέχουν λειτουργικά διαφορετικές πρωτεΐνες: ενζυμικές πρωτεΐνες, πρωτεΐνες που δρουν ως αντλίες, φορείς, κανάλια ιόντων, ρυθμιστικές πρωτεΐνες και δομικές πρωτεΐνες (Εικόνα 9).
Εάν τέτοια εξειδικευμένα σύμπλοκα πρωτεϊνών:
Βυθισμένες στη λιπιδική φάση και συγκρατημένες από υδρόφοβους δεσμούς (λιποπρωτεΐνες), ονομάζονται «ενσωματωμένες πρωτεΐνες».
συγκρατούνται στην εσωτερική και εξωτερική επιφάνεια των μεμβρανών με ηλεκτροστατικούς δεσμούς (υδρόφιλες πρωτεΐνες), αλληλεπιδρώντας με τις υδρόφιλες κεφαλές των πολικών λιπιδίων, ονομάζονται «περιφερικές πρωτεΐνες».
Οι περιφερειακές πρωτεΐνες συνδέονται με τη μεμβράνη με την προσκόλληση σε ενσωματωμένες πρωτεΐνες ή τη λιπιδική διπλοστιβάδα με ασθενείς δεσμούς: υδρογόνο, ηλεκτροστατική, γέφυρες αλατιού. Είναι κυρίως διαλυτά στο νερό και διαχωρίζονται εύκολα από τη μεμβράνη χωρίς να την καταστρέφουν. Αρκετές περιφερειακές πρωτεΐνες παρέχουν μια σύνδεση μεταξύ των μεμβρανών και του κυτταροσκελετού.
Οι ενσωματωμένες πρωτεΐνες μεμβράνης είναι αδιάλυτες στο νερό. Η θέση των πρωτεϊνών στη μεμβράνη επηρεάζεται από τη σύνθεση των φωσφολιπιδίων (ελεύθερων και δεσμευμένων), το μέγεθος του ηλεκτροστατικού φορτίου της μεμβράνης.
Πρόσφατα, έχει αποδειχθεί η ύπαρξη μιας τρίτης ομάδας πρωτεϊνών, των λεγόμενων «αγκυροβολημένων» πρωτεϊνών στη μεμβράνη. Αυτές οι πρωτεΐνες στερεώνονται στη μεμβράνη από ένα ειδικό μόριο, το οποίο μπορεί να είναι ένα λιπαρό οξύ, στερόλη, ισοπρενοειδή ή φωσφατιδυλινοσιτόλη.
Ένα χαρακτηριστικό της λιπιδικής σύνθεσης του πλάσματος σε σύγκριση με άλλες φυτικές κυτταρικές μεμβράνες είναι η υψηλή περιεκτικότητα σε στερόλες, αλλά σε αντίθεση με την πλασματική μεμβράνη ενός ζωικού κυττάρου, το πλασμάλεμα χαρακτηρίζεται από υψηλή μεταβλητότητα της σύνθεσής τους ανάλογα με τον τύπο του φυτού. όργανο και ιστό.
Οι φορείς είναι συγκεκριμένες πρωτεΐνες που μπορούν να συνδεθούν με μια
.
Οι αντλίες (αντλίες) είναι αναπόσπαστες πρωτεΐνες μεταφοράς που παρέχουν ενεργά ιόντα. Ο όρος «αντλία» υποδηλώνει ότι η ροή είναι με την κατανάλωση ελεύθερης ενέργειας και αντίθετα με την ηλεκτροχημική κλίση. Η ενέργεια που χρησιμοποιείται για την ενεργό είσοδο ιόντων παρέχεται από τις διαδικασίες της αναπνοής και της φωτοσύνθεσης και συσσωρεύεται κυρίως στο ATP. Όπως γνωρίζετε, για να χρησιμοποιηθεί η ενέργεια που περιέχεται στο ATP, αυτή η ένωση πρέπει να υδρολυθεί σύμφωνα με την εξίσωση ATP + HOH → ADP + Ph n . Τα ένζυμα που υδρολύουν το ATP ονομάζονται τριφωσφατάσες αδενοσίνης (ATPases). Διάφορες ΑΤΡάσες βρέθηκαν σε κυτταρικές μεμβράνες: K + /Na + -ATPase; Ca 2+ - ΑΤΡάση; H + -ATPase. H + –ATPase (H +-αντλία ή αντλία υδρογόνου) είναι ο κύριος μηχανισμός ενεργού μεταφοράς στα κύτταρα των φυτών, των μυκήτων και των βακτηρίων. Η H + - ATPase λειτουργεί στο πλάσμα και εξασφαλίζει την απελευθέρωση πρωτονίων από το κύτταρο, η οποία οδηγεί στο σχηματισμό ηλεκτροχημικής διαφοράς δυναμικού στη μεμβράνη. Η H + -ATPase μεταφέρει πρωτόνια στην κοιλότητα του κενοτοπίου και στις δεξαμενές της συσκευής Golgi.
Ο υπολογισμός δείχνει ότι για να διαχέεται 1 mol άλατος έναντι της βαθμίδας συγκέντρωσης, είναι απαραίτητο να δαπανηθούν περίπου 4600 J. Ταυτόχρονα, απελευθερώνονται 30660 J/mol κατά την υδρόλυση ATP. Επομένως, αυτή η ενέργεια ATP θα πρέπει να είναι αρκετή για να μεταφέρει μερικά mole αλατιού. Υπάρχουν στοιχεία που δείχνουν μια ευθέως αναλογική σχέση μεταξύ της δραστηριότητας της ΑΤΡάσης και της πρόσληψης ιόντων. Η ανάγκη για μόρια ΑΤΡ να πραγματοποιήσουν τη μεταφορά επιβεβαιώνεται επίσης από το γεγονός ότι οι αναστολείς που διαταράσσουν τη συσσώρευση αναπνευστικής ενέργειας στο ΑΤΡ (παραβίαση της σύζευξης οξείδωσης και φωσφορυλίωσης), ιδιαίτερα η δινιτροφαινόλη, αναστέλλουν τη ροή των ιόντων.
Οι αντλίες χωρίζονται σε δύο ομάδες:
1) ηλεκτρογονικά, τα οποία πραγματοποιούν ενεργή μεταφορά ενός ιόντος οποιουδήποτε φορτίου προς μία μόνο κατεύθυνση. Αυτή η διαδικασία οδηγεί στη συσσώρευση ενός τύπου φορτίου στη μία πλευρά της μεμβράνης.
2) ηλεκτρικά ουδέτερο, στο οποίο η μεταφορά ενός ιόντος προς μία κατεύθυνση συνοδεύεται από κίνηση ιόντος του ίδιου σημείου προς την αντίθετη κατεύθυνση ή από τη μεταφορά δύο ιόντων με φορτία ίδιου μεγέθους, αλλά διαφορετικού πρόσημου, προς την ίδια κατεύθυνση.
Προσδιορισμός προσρόφησης
Το φάσμα των εκδηλώσεων της τοξικής διαδικασίας καθορίζεται από τη δομή του ξενοβιοτικού. Για να δηλώσει την ποσότητα μιας ουσίας που δρα σε ένα βιολογικό αντικείμενο, χρησιμοποιείται η έννοια της «δόσης». Η εξάρτηση «δόσης-αποτελέσματος» μπορεί να εντοπιστεί σε όλα τα επίπεδα οργάνωσης της ζωντανής ύλης: από το μοριακό έως τον πληθυσμό. Σε αυτή την περίπτωση, στη συντριπτική πλειονότητα των περιπτώσεων, θα καταγραφεί ένα γενικό πρότυπο: με αύξηση της δόσης, ο βαθμός βλάβης στο σύστημα αυξάνεται. Ένας αυξανόμενος αριθμός των συστατικών του στοιχείων εμπλέκεται στη διαδικασία. Το απλούστερο αντικείμενο που είναι απαραίτητο για την καταγραφή της βιολογικής δράσης ενός ξενοβιοτικού είναι ένα κύτταρο.
Πολλά ξενοβιοτικά δρουν άμεσα με την προσρόφησή τους στην κυτταρική επιφάνεια (μεμβράνη). Η προσροφητική επιφάνεια σε ένα στοιχείο μπορεί να υπερβαίνει τον όγκο κατά πολλές τάξεις μεγέθους. Οι πρωτεΐνες και τα μεγάλα μόρια σε διάλυμα βρίσκονται σε κολλοειδή κατάσταση και παρέχουν μια τεράστια επιφάνεια για προσρόφηση. Λαμβάνοντας υπόψη το φαινόμενο της προσρόφησης, θα πρέπει να ληφθούν υπόψη:
1) τα φυσικοχημικά χαρακτηριστικά των ουσιών μετά την προσρόφησή τους διαφέρουν από τις ιδιότητές τους στο διάλυμα.
2) η ουσία μπορεί να απορροφηθεί αναστρέψιμα στην επιφάνεια του κυττάρου.
3) η προσρόφηση καθορίζεται από το άθροισμα όλων των χημικών δεσμών που σχηματίζονται μεταξύ των μορίων ή των μορίων και της επιφάνειας.
4) η διαδικασία της προσρόφησης οφείλεται στους ίδιους τύπους δεσμών (ειδικά van der Waal, υδρογόνο και ιοντικό) με τις χημικές αντιδράσεις που συμβαίνουν σε ολόκληρο τον όγκο της ουσίας.
5) χαρακτηριστικά που καθορίζουν τις ποσοτικές διαφορές μεταξύ των αντιδράσεων που συμβαίνουν στην επιφάνεια και στο διάλυμα:
α) Δημιουργείται 100% συγκέντρωση της ουσίας στην επιφάνεια. Δεδομένου ότι η προσροφημένη ουσία έχει αμελητέα διαλυτότητα, τότε σε μια τέτοια συγκέντρωση, η πιθανότητα χημικής αλληλεπίδρασης αυξάνεται σημαντικά.
β) η επιφάνεια χαρακτηρίζεται από την παρουσία ακόρεστων σθένων, τα οποία σε ένα στερεό δαπανώνται για τη σύνδεση μεταξύ τους των συστατικών του ατόμων.
γ) ένα μόριο που προσροφάται από ένα διάλυμα στην επιφάνεια υπόκειται σε δυνάμεις που τείνουν να το επαναφέρουν στο διάλυμα. Ένα μέτρο της ικανότητας μιας ουσίας να επιστρέφει στο περιβάλλον, δηλαδή να απορροφάται, είναι η διαλυτότητά της.
Η προσρόφηση χωρίζεται σε μη ειδική και ειδική.
Λειτουργίες μεμβράνης
1) Δομική λειτουργία. Ένα ευκαρυωτικό κύτταρο, σε αντίθεση με ένα προκαρυωτικό κύτταρο, έχει πολλά εσωτερικά διαμερίσματα (διαμερίσματα) που περιβάλλονται από μεμβράνες και διαφέρουν ως προς τον βαθμό δραστηριότητας των χημικών ενώσεων που περιέχονται σε αυτές και τα συστήματα που ρυθμίζουν τους μετασχηματισμούς τους. Τα διαμερίσματα ενός ευκαρυωτικού κυττάρου ονομάζονται οργανίδια. Μονομεμβρανικά δομικά συστατικά των κυττάρων: πλασμάλημμα, ενδοπλασματικό δίκτυο, σύμπλεγμα Golgi, λυσοσώματα, κενοτόπιο.
Δομικά συστατικά δύο μεμβρανών κυττάρων: πυρήνας, μιτοχόνδρια, πλαστίδια.
2) Λειτουργία φραγμού . Για τα κύτταρα και τα υποκυτταρικά σωματίδια, οι μεμβράνες χρησιμεύουν ως μηχανικό φράγμα που τα χωρίζει από τον εξωτερικό χώρο.
3) Έλεγχος απορρόφησης και έκκρισης ουσιών (λειτουργία μεταφοράς). Η πλασματική μεμβράνη περιέχει πολλά συστήματα μεταφοράς, κυρίως για τη μεταφορά ιόντων - διαύλων ιόντων, φορέων ιόντων και αντλιών ιόντων. Χάρη σε αυτά, πραγματοποιείται μια πολύ ακριβής και επιλεκτική μεταφορά ιόντων τόσο μέσα στο κύτταρο όσο και έξω, δηλ. βέλτιστη παροχή των κυττάρων με τα απαραίτητα ιόντα.
4) Συσσώρευση και μετατροπή ενέργειας. Το πλάσμα οποιουδήποτε κυττάρου είναι μια ενεργοποιημένη μεμβράνη, δηλαδή υπάρχει μια βαθμίδα ηλεκτροχημικού δυναμικού Δμ H+ , η οποία χρησιμοποιείται για την εκτέλεση χρήσιμης εργασίας, κυρίως για την ενεργό μεταφορά ουσιών μέσω της μεμβράνης.
5) Τοποθέτηση και διασφάλιση της εργασίας των ενζύμων. Η πλασματική μεμβράνη περιέχει πολλά ένζυμα. Αυτά είναι ένζυμα κατασκευής κυτταρικού τοιχώματος (συνθάση κυτταρίνης), ένζυμα συστημάτων σηματοδότησης (φωσφολιπάσες C, Α, αδενυλική κυκλάση)
6) Λειτουργία υποδοχέα. Το κυτταρικό πλάσμα είναι ένα μωσαϊκό από διάφορους υποδοχείς για ενδογενή σήματα (κυρίως φυτοορμόνες) και εξωτερικές επιδράσεις. Η θερμοκρασία αλλάζει την ακαμψία (ρευστότητα) της μεμβράνης, δηλ. τις μηχανικές του ιδιότητες. Μια αλλαγή σε αυτές τις ιδιότητες της πλασματικής μεμβράνης οδηγεί στο άνοιγμα ή το κλείσιμο των διαύλων μηχανοαισθητήρων ιόντων που βρίσκονται σε αυτήν.
7) Λειτουργία σήματος. Πολλά συστατικά του πλάσματος μετά την αντίληψη των σημάτων χρησιμεύουν ως πηγή δευτερευόντων αγγελιοφόρων - μορίων που «μεταδίδουν» το σήμα κατά μήκος της κούρσας αναμετάδοσης και το ενισχύουν.
Το κυτταρικό τοίχωμα παρέχει μηχανική αντοχή στο στοιχείο, δίνοντάς του μια άκαμπτη (άκαμπτη) δομή, έτσι ώστε το κύτταρο να μπορεί να αντέξει υψηλή εσωτερική οσμωτική πίεση (5-20 MPa). Επιπλέον, το κυτταρικό τοίχωμα μπορεί να παρέχει κάποιο βαθμό επιλεκτικής διαπερατότητας σε ουσίες μικρού μοριακού βάρους, καθώς και την ικανότητα να αλληλεπιδρά με άλλα κύτταρα, ιούς και φυσικές επιφάνειες. Η δομή του κυτταρικού τοιχώματος σε διαφορετικούς οργανισμούς έχει τα δικά της χαρακτηριστικά.
Τα κύτταρα των περισσότερων ιστών των πολυκύτταρων ζώων δεν περιέχουν έντονο κυτταρικό τοίχωμα. Τα φυτικά κύτταρα, αντίθετα, έχουν ένα πολύ περίπλοκο κυτταρικό τοίχωμα κατασκευασμένο από μικροϊνίδια κυτταρίνης ενσωματωμένα σε μια μήτρα (πηκτίνης και ημικυτταρινών).
Τα κυτταρικά τοιχώματα των ζυμομυκήτων και των νηματοειδών μυκήτων αποτελούνται από ομο- και ετεροπολυσακχαρίτες (γλυκάνες, χιτίνη) και ένα σύμπλεγμα πρωτεϊνών.Το πάχος αυτών των στρωμάτων φτάνει το 1 μm.
Το κυτταρικό τοίχωμα των θετικών στον κεραυνό βακτηρίων είναι κατασκευασμένο κυρίως από γέφυρες αμινοξέων που περιέχουν ετεροπολυσακχαρίτη μουρεΐνης. Στα gram-αρνητικά βακτήρια, το στρώμα μουρεΐνης είναι μικρό, αλλά το κυτταρικό τοίχωμα περιέχει μια εξωτερική μεμβράνη φτιαγμένη από φωσφολιπίδια, πρωτεΐνες και λιποπολυσακχαρίτη, η οποία παρέχει κάποιο βαθμό επιλεκτικής διαπερατότητας και περιέχει υποδοχείς φάγου και αντιγόνα. Το πάχος του κυτταρικού τοιχώματος κυμαίνεται από 15 έως 80 nm
Εξάρτηση από τη θερμοκρασία του υδρόφοβου πάχους dL των διπλών στοιβάδων που σχηματίζονται από (1) διπαλμιτοϋλφωσφατιδυλοχολίνη και (2) διμυριστοϋλφωσφατιδυλοχολίνη.
https://www.chem.msu.ru/rus/teaching/kolman/212.htm
Οι μιτοχονδριακές μεμβράνες περιέχουν ενσωματωμένες μεμβρανικές πρωτεΐνες. Η εξωτερική μεμβράνη περιλαμβάνει πορίνες , οι οποίες σχηματίζουν πόρους και καθιστούν τις μεμβράνες διαπερατές σε ουσίες με μοριακό βάρος έως 10 kDa (βλ. εικ. 223 ). Η εσωτερική μεμβράνη των μιτοχονδρίων είναι αδιαπέραστη από τα περισσότερα μόρια. η εξαίρεση είναι το O 2 , CO 2 , H 2 0. Η εσωτερική μεμβράνη των μιτοχονδρίων χαρακτηρίζεται από ασυνήθιστα υψηλή περιεκτικότητα σε πρωτεΐνες (75%). Αυτές περιλαμβάνουν πρωτεΐνες-φορείς μεταφοράς (βλέπε εικ. 215 ), ένζυμα, συστατικά της αναπνευστικής αλυσίδας και συνθάση ATP . Επιπλέον, περιέχει ένα ασυνήθιστο φωσφολιπίδιο που ονομάζεται καρδιολιπίνη.(βλ. σελ. 56 ). Η μήτρα είναι επίσης εμπλουτισμένη σε πρωτεΐνες, ιδιαίτερα σε ένζυμα κύκλου κιτρικών.
Τα μιτοχόνδρια αποτελούν το «ηλεκτρικό κέντρο» του κυττάρου, αφού λόγω της οξειδωτικής αποικοδόμησης των θρεπτικών συστατικών, συνθέτουν το μεγαλύτερο μέρος του ATP (ATP) που χρειάζεται το κύτταρο. Οι ακόλουθες μεταβολικές διεργασίες εντοπίζονται στα μιτοχόνδρια: μετατροπή πυροσταφυλικού σε ακετυλο-CoA που καταλύεται από το σύμπλοκο πυροσταφυλικής αφυδρογονάσης: κύκλος κιτρικού. αναπνευστική αλυσίδα , που σχετίζεται με τη σύνθεση του ATP (ο συνδυασμός αυτών των διεργασιών ονομάζεται " οξειδωτική φωσφορυλίωση "). διάσπαση λιπαρών οξέων με β-οξείδωση και εν μέρει τον κύκλο της ουρίας . Τα μιτοχόνδρια τροφοδοτούν επίσης το κύτταρο με προϊόντα ενδιάμεσου μεταβολισμού και λειτουργούν, μαζί με το ER, ως αποθήκη ιόντων ασβεστίου., η οποία με τη βοήθεια αντλιών ιόντων διατηρεί τη συγκέντρωση του Ca 2+ στο κυτταρόπλασμα σε σταθερά χαμηλό επίπεδο (κάτω από 1 μmol/l).
Η κύρια λειτουργία των μιτοχονδρίων είναι η σύλληψη πλούσιων σε ενέργεια υποστρωμάτων (λιπαρά οξέα, πυροσταφυλικό, ανθρακικός σκελετός αμινοξέων) από το κυτταρόπλασμα και η οξειδωτική διάσπασή τους με το σχηματισμό CO 2 και H 2 O, σε συνδυασμό με τη σύνθεση του ATP.
Οι αντιδράσεις του κύκλου των κιτρικών οδηγούν στην πλήρη οξείδωση των ενώσεων που περιέχουν άνθρακα (CO 2 ) και στο σχηματισμό αναγωγικών ισοδυνάμων, κυρίως με τη μορφή ανηγμένων συνενζύμων. Οι περισσότερες από αυτές τις διαδικασίες λαμβάνουν χώρα στη μήτρα. Τα ένζυμα της αναπνευστικής αλυσίδας που επαναοξειδώνουν ανηγμένα συνένζυμα βρίσκονται στην εσωτερική μιτοχονδριακή μεμβράνη. Το NADH και το δεσμευμένο σε ένζυμο FADH 2 χρησιμοποιούνται ως δότες ηλεκτρονίων για τη μείωση του οξυγόνου και το σχηματισμό νερού . Αυτή η εξαιρετικά εξεργολογική αντίδραση είναι πολλαπλών σταδίων και σχετίζεται με τη μεταφορά πρωτονίων (H + ) μέσω της εσωτερικής μεμβράνης από τη μήτρα στον διαμεμβρανικό χώρο (βλ. Εικ. 143). Ως αποτέλεσμα, δημιουργείται μια ηλεκτροχημική κλίση στην εσωτερική μεμβράνη (βλ. Εικ. 129 ). Στα μιτοχόνδρια, η ηλεκτροχημική βαθμίδα χρησιμοποιείται για τη σύνθεση ATP από ADP (ADP) και ανόργανο φωσφορικό (Pi ) που καταλύεται από συνθάση ATP . Η ηλεκτροχημική κλίση είναι επίσης η κινητήρια δύναμη πίσω από έναν αριθμό συστημάτων μεταφοράς (βλ. Εικ. 215 )
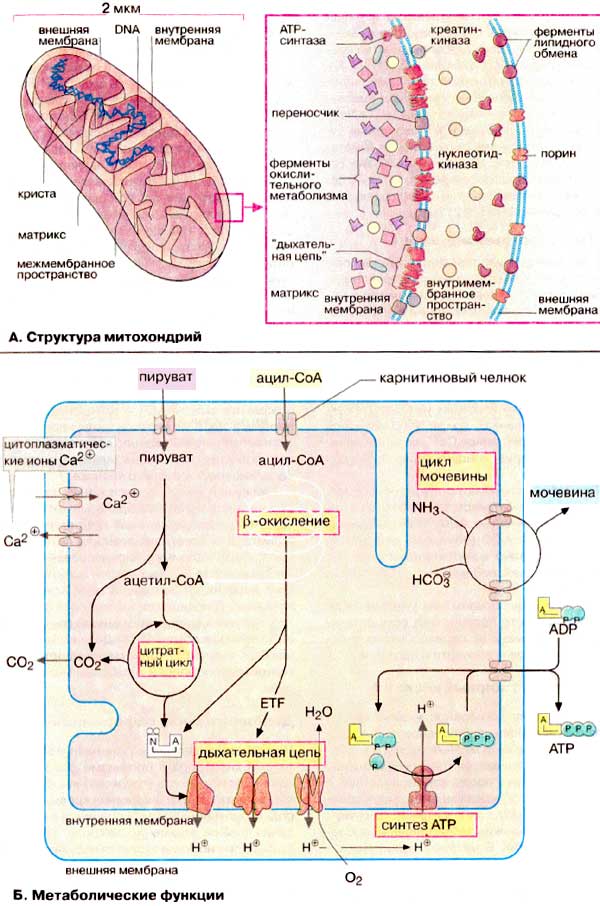 Α. Δομή μιτοχονδρίων
Α. Δομή μιτοχονδρίων
Τα μιτοχόνδρια είναι οργανίδια μεγέθους βακτηρίου (περίπου 1 x 2 μm). Βρίσκονται σε μεγάλους αριθμούς σχεδόν σε όλα τα ευκαρυωτικά κύτταρα. Τυπικά, ένα κύτταρο περιέχει περίπου 2000 μιτοχόνδρια, ο συνολικός όγκος των οποίων είναι έως και το 25% του συνολικού όγκου των κυττάρων. Το μιτοχόνδριο περιορίζεται από δύο μεμβράνες - μια λεία εξωτερική και μια διπλωμένη εσωτερική , η οποία έχει πολύ μεγάλη επιφάνεια. Οι πτυχές της εσωτερικής μεμβράνης εισέρχονται βαθιά στη μιτοχονδριακή μήτρα, σχηματίζοντας εγκάρσια διαφράγματα . Ο χώρος μεταξύ της εξωτερικής και της εσωτερικής μεμβράνης αναφέρεται συνήθως ως ο ενδομεμβρανικός χώρος .
Οι διαφορετικοί τύποι κυττάρων διαφέρουν μεταξύ τους τόσο στον αριθμό και το σχήμα των μιτοχονδρίων όσο και στον αριθμό των κριστών. Ειδικά πολλά cristae έχουν μιτοχόνδρια σε ιστούς με ενεργές οξειδωτικές διεργασίες, για παράδειγμα, στον καρδιακό μυ. Παραλλαγές στο σχήμα των μιτοχονδρίων, που εξαρτάται από τη λειτουργική τους κατάσταση, μπορούν επίσης να παρατηρηθούν σε ιστούς του ίδιου τύπου. Τα μιτοχόνδρια είναι εύκαμπτα και εύκαμπτα οργανίδια.
Οι μιτοχονδριακές μεμβράνες περιέχουν ενσωματωμένες μεμβρανΟι αντιδράσεις του κύκλου των κιτρικών οδηγούν στην πλήρη οξείδωση των ενώσεων που περιέχουν άνθρακα (CO 2 ) και στο σχηματισμό αναγωγικών ισοδυνάμων, κυρίως με τη μορφή ανηγμένων συνενζύμων. Οι περισσότερες από αυτές τις διαδικασίες λαμβάνουν χώρα στη μήτρα. Τα ένζυμα της αναπνευστικής αλυσίδας που επαναοξειδώνουν ανηγμένα συνένζυμα βρίσκονται στην εσωτερική μιτοχονδριακή μεμβράνη. Το NADH και το δεσμευμένο σε ένζυμο FADH 2 χρησιμοποιούνται ως δότες ηλεκτρονίων για τη μείωση του οξυγόνου και το σχηματισμό νερού . Αυτή η εξαιρετικά εξεργολογική αντίδραση είναι πολλαπλών σταδίων και σχετίζεται με τη μεταφορά πρωτονίων (H + ) μέσω της εσωτερικής μεμβράνης από τη μήτρα στον διαμεμβρανικό χώρο (βλ. Εικ. 143). Ως αποτέλεσμα, δημιουργείται μια ηλεκτροχημική κλίση στην εσωτερική μεμβράνη (βλ. Εικ. 129 ). Στα μιτοχόνδρια, η ηλεκτροχημική βαθμίδα χρησιμοποιείται για τη σύνθεση ATP από ADP (ADP) και ανόργανο φωσφορικό (Pi ) που καταλύεται από συνθάση ATP . Η ηλεκτροχημική κλίση είναι επίσης η κινητήρια δύναμη πίσω από έναν αριθμό συστημάτων μεταφοράς (βλ. Εικ. 215 ).
Физиология клеточных мембран.pdf - christmedschool.com
Физиология клеточных мембран.pdf - christmedschool.com



Οι χλωροπλάστες των χερσαίων φυτών έχουν διάμετρο περίπου 4 έως 8 μm. Ωστόσο, σε πολλά φύκια με μόνο έναν μόνο χλωροπλάστη ανά κύτταρο, μπορεί να καταλάβει μεγάλο μέρος του κυττάρου.
Η δομική σύνθεση των χλωροπλαστών είναι παρόμοια με αυτή των κυανοβακτηρίων (γαλαζοπράσινα βακτήρια, παλαιότερα γνωστά ως γαλαζοπράσινα φύκια). Οι χλωροπλάστες (σχεδόν πάντα) έχουν το δικό τους πλαστιδικό DNA (Chloroplast DNA, συντομευμένο cpDNA ή ctDNA) μαζί με τα δικά τους ριβοσώματα (πλασιδικά ριβοσώματα ή πλαστοριβοσώματα) και είναι παρόμοια με τα μιτοχόνδρια.[1][2] Το γονιδίωμα του χλωροπλάστη και των άλλων πλαστιδίων ονομάζεται επίσης πλάστωμα.
Οι χλωροπλάστες καλύπτονται από δύο βιομεμβράνες, μέσα στις οποίες βρίσκεται το στρώμα ως πλασματική φάση. Το στρώμα, με τη σειρά του, διαπερνάται από θυλακοειδή μεμβράνες, παράγωγα της εσωτερικής μεμβράνης. Με την εξαίρεση πολλών φωτοτροφικών πρωτιστών, οι χλωροπλάστες των υψηλότερων φωτοτροφών έχουν επίπεδες, στρογγυλές προεξοχές αυτών των μεμβρανών που υπερτίθενται «σαν ρολό χρήματος» σε πολλά σημεία – μια τέτοια στοίβα θυλακοειδή ονομάζεται granum (pl. grana). Στις μεμβράνες των θυλακοειδών αποθηκεύονται διάφορες χρωστικές, κυρίως η πράσινη χρωστική χλωροφύλλη. Ιδιαίτερα μεγάλη ποσότητα της εντοπίζεται στις μεμβράνες της grana, γι' αυτό και εμφανίζονται έντονο πράσινο χρώμα.
Οι χρωστικές μπορούν να απορροφήσουν φως ορισμένων μηκών κύματος και η απορροφούμενη ενέργεια χρησιμοποιείται για την παραγωγή ATP από ADP και φωσφορικά (βλ. φωτοτροφία). Το ATP χρησιμεύει ως φορέας ενέργειας για τη δημιουργία γλυκόζης ή αμύλου από το CO2 και το νερό.
Κατασκευή δυναμικού χημειοσμωτικής μεμβράνης
Η βιογένεση αυτών των τριών μεμβρανικών συστημάτων εξηγεί το γεγονός ότι το δυναμικό της μεμβράνης δημιουργείται από μια βαθμίδα πρωτονίου κατά μήκος της μεμβράνης του θυλακοειδούς στους χλωροπλάστες (το εσωτερικό του θυλακοειδούς έχει όξινο περιβάλλον), ενώ στα μιτοχόνδρια ο διαμεμβρανικός χώρος (περιοχή μεταξύ της εσωτερικής και της εξωτερικής μεμβράνης ) συνδέεται χημειοσμωτικά με ιόντα Η+ είναι φορτισμένα. Ομοίως, η συνθάση ATP (γνωστή και ως FoF1-ATPase) είναι ένα ένζυμο ενσωματωμένο στη θυλακοειδή μεμβράνη στους χλωροπλάστες (το τμήμα CF1 προεξέχει στο στρώμα) και στα μιτοχόνδρια είναι ένα συστατικό της εσωτερικής μεμβράνης (τμήμα F1 που βλέπει στη μήτρα). Έτσι, και στα δύο συστήματα, το ATP παραδίδεται στη μήτρα/στρώμα. Σε αντάλλαγμα για το ADP, μπορεί να εισέλθει στο κυτταρόπλασμα του κυττάρου.
https://de.wikipedia.org/wiki/Chloroplast
Σήμερα γνωρίζουμε ότι τα βακτήρια έχουν επίσης έναν κυτταροσκελετό του οποίου οι πρωτεΐνες σχετίζονται εξελικτικά με αυτές του ευκαρυωτικού κυτταροσκελετού. Είναι γνωστό από πειράματα στα βρύα φύλλων Physcomitrella patens (συμπεριλαμβανομένων των βρύων νοκ-άουτ) ότι οι πρωτεΐνες FtsZ, τα ομόλογα τουμπουλίνης, όχι μόνο προκαλούν τη διαίρεση των χλωροπλαστών,[3] αλλά μπορούν επίσης να σχηματίσουν ένα σύνθετο δίκτυο στους χλωροπλάστες. Δεδομένου ότι αυτά τα δίκτυα θυμίζουν έντονα τον κυτταροσκελετό, ο Ralf Reski επινόησε τον όρο "πλαστοσκελετός" για αυτές τις πολύπλοκες δομές το 2000 και υπέθεσε ότι εκπληρώνουν παρόμοιες λειτουργίες στα πλαστίδια όπως ο κυτταροσκελετός για ολόκληρο το κύτταρο.[4][5]
DNA χλωροπλάστη
→ Κύριο άρθρο: DNA χλωροπλάστη
Το DNA του χλωροπλάστη ανιχνεύθηκε για πρώτη φορά το 1962,[6] και η αλληλουχία ενός πλαστόμου έγινε για πρώτη φορά το 1986.[7] Από τότε, εκατοντάδες DNA χλωροπλαστών από διαφορετικά είδη έχουν αναλυθεί σε αλληλουχία. Στις περισσότερες περιπτώσεις όμως πρόκειται για χλωροπλάστες, δηλ. H. φυτά της γης ή πράσινα φύκια. Τα γλαυκόφυτα (Glaucophyta), τα κόκκινα φύκια (Rhodophyta syn. Rhodophyceae) και άλλες ομάδες φυκών υποεκπροσωπούνται σε μεγάλο βαθμό.[8] Το DNA των χλωροπλαστών -τουλάχιστον στους χλωροπλάστες (πράσινα φύκια και φυτά της γης) - είναι συνήθως δομημένο σε έναν δακτύλιο όπως στα περισσότερα βακτήρια. Το γονιδίωμα του χλωροπλάστη (πλαστόμ) έχει τμήματα που υπάρχουν ως ανεστραμμένες επαναλήψεις, παρόμοια με το γονιδίωμα των κυανοβακτηρίων. Στα μιτοχόνδρια και στα πλαστίδια όπως οι χλωροπλάστες, όπως και στα βακτήρια, το DNA συνήθως συμπυκνώνεται σε νουκλεοειδή (ισοδύναμα πυρήνων). Αυτό εξασφαλίζεται από τις λεγόμενες πρωτεΐνες τύπου ιστόνης (HLPs). Αυτές είναι ομόλογες μεταξύ τους, αλλά μόνο λειτουργικά παρόμοιες (ανάλογες) με τις αληθινές ιστόνες στον κυτταρικό πυρήνα των ευκαρυωτικών κυττάρων (ευκυττάρων). Τα γονίδια που κωδικοποιούν την πρωτεΐνη μεταγράφονται σε αγγελιοφόρο RNA, το οποίο χρησιμεύει ως πρότυπο για τη σύνθεση πρωτεϊνών (μετάφραση) στα ριβοσώματα του ίδιου του σώματος (πλαστοριβοσώματα). Αυτά τα πλαστιδικά ριβοσώματα είναι παρόμοια σε σύσταση και δομή με εκείνα των βακτηρίων και των μιτοχονδρίων, αλλά είναι σημαντικά μικρότερα και απλούστερα στη δομή από εκείνα των γύρω ευκυττάρων.
Προέλευση των χλωροπλαστών - θεωρία ενδοσυμβίωσης
→ Κύριο άρθρο: Θεωρία Ενδοσυμβίωσης
Ο Γερμανός βοτανολόγος Andreas Franz Wilhelm Schimper είχε ήδη διαπιστώσει το 1883 ότι οι χλωροπλάστες είναι πολύ παρόμοιοι με τα κυανοβακτήρια και έτσι υπέδειξε ήδη μια συμβιωτική προέλευση των χλωροπλαστών, έτσι ώστε να μπορεί να θεωρηθεί ο πρωτοπόρος της μεταγενέστερης διατυπωμένης θεωρίας του ενδοσυμβίου.[9] Παρόμοιες ιδέες διατυπώθηκαν εκείνη την εποχή από τους Sachs και Altmann.[10] Ο Ρώσος βιολόγος Konstantin Sergejewitsch Mereschkowski υιοθέτησε αυτή την ιδέα το 1905[11] και ο Ivan Wallin το 1922[12][13] και την έκανε πιο συγκεκριμένη.
Τόσο οι χλωροπλάστες όσο και τα κυανοβακτήρια έχουν διπλή μεμβράνη, DNA, ριβοσώματα και θυλακοειδή. Εμφανίζονται εξιδανικευμένες εκδοχές ενός χλωροπλάστου ενός ανώτερου φυτού (αγγειακό φυτό) και ενός κυανοβακτηρίου - υπάρχει μεγάλη ποικιλομορφία τόσο μεταξύ των χλωροπλαστών όσο και των κυανοβακτηρίων. Το κυτταρόπλασμα που βρίσκεται μέσα στα βακτήρια είναι ομόλογο με το στρώμα των χλωροπλαστών, καθώς και με τη μήτρα των μιτοχονδρίων.
Το πώς οι χλωροπλάστες διαιρούνται και αλλάζουν σχήμα ήταν άγνωστο για πολύ καιρό. Δεδομένου ότι δεν ήταν δυνατό να καλλιεργηθούν απομονωμένοι χλωροπλάστες in vitro (στο εργαστήριο σε μέσα καλλιέργειας), η θεωρία μπόρεσε να καθιερωθεί μόνο στις αρχές της δεκαετίας του 1970 (Lynn Margulis), όταν το DNA ανιχνεύθηκε και στους δύο τύπους οργανιδίων.[14][14][14] 15] Ο ανεξάρτητος «πολλαπλασιασμός» των χλωροπλαστών, παρόμοιος με αυτόν των βακτηριδίων, χωρίς δομική σύζευξη με την κυτταρική διαίρεση του περιβάλλοντος ευκαρυωτικού κυττάρου, θεωρείται ένα επιπλέον επιχείρημα υπέρ της θεωρίας του ενδοσυμβίωσης.
Πρωτοπαθής ενδοσυμβίωση
Δ.Σ.
https://science.fandom.com/el/wiki/%CE%9C%CE%B9%CE%BA%CF%81%CE%BF%CE%BA%CE%B1%CE%BD%CE%BF%CE%BD%CE%B9%CE%BA%CE%AE_%CE%A3%CF%85%CE%BB%CE%BB%CE%BF%CE%B3%CE%AE
https://users.auth.gr/vinga/courses/statistical/chapters/chaptersa-11.pdf
https://www.studocu.com/gr/document/eoniko-metsobio-polytexneio/maths-3/fermions-and-bosons/43431326
Αλλαγή στην τιμή μιας παραμέτρου, αντιστρέφει το αποτέλεσμα
Σε χαμηλές συγκεντρώσεις η αδρεναλίνη επιδρά μόνο στους β υποδοχείς (χαλαρή αγειοδιαστολή), διαφορετικά στους α υποδοχείς (αγγειοσυστολή)
Στο δέρμα και στον γαστρεντερικό σωλήνα, η αδρεναλίνη προκαλεί αγγειοσυστολή , ενώ στους μυς σε μικρή συγκέντρωση αγγειοδιαστολή, σε μεγάλη αγγειοσυστολή
Το ασβέστιο είναι ο συνδετικός κρίκος μεταξύ της διέγερσης και της συστολής. Οι σκελετικοί μύες διεγείρονται προς συστολή από την απελευθέρωση της ακετυλοχολίνης (ACh) στις νευρομυϊκές συνάψεις, ανάμεσα στα τελικά κομβία των κινητικών νευρώνων και στις μυϊκές ίνες. Θυμηθείτε ότι η πρόσδεση της ACh στην τελική κινητική πλάκα μιας μυϊκής ίνας επάγει μεταβολές στη διαπερατότητα της κυτταρικής μεμβράνης της μυϊκής ίνας, γεγονός που οδηγεί στη γένεση ενός δυναμικού ενέργειας το οποίο διαδίδεται σε όλη την επιφάνεια της κυτταρικής μεμβράνης του μυϊκού κυττάρου (βλ. Ενότητα 7.3).
Δύο μεμβρανικές δομές μέσα στη μυϊκή ίνα διαδραματίζουν σημαντικό ρόλο στη σύζευξη της διέγερσης με τη συστολή: τα εγκάρσια σωληνάρια και το σαρκοπλασματικό δίκτυο. Στη συνέχεια θα εξετάσουμε τη δομή και τη λειτουργία καθενός από αυτά.
Η εξωτερική μεμβράνη χρησιμεύει κυρίως ως φράγμα διάχυσης για μεγάλα μόρια. Όπως ένα «φίλτρο» εμποδίζει τα βιοπολυμερή να εισέλθουν στον περιπλασμικό χώρο, αλλά κυρίως εμποδίζει τη διαφυγή πρωτεϊνών και ενζύμων από το περίπλασμα στο περιβάλλον. Η μεμβράνη περιέχει πολυάριθμες πορίνες και άλλες αναπόσπαστες πρωτεΐνες που είναι απαραίτητες για τις διαδικασίες μεταφοράς ή τη λήψη σημάτων και περιβαλλοντικών ερεθισμάτων. Οι δομές β-βαρελιού σε σχήμα βαρελιού είναι χαρακτηριστικές των ενσωματωμένων πρωτεϊνών της εξωτερικής μεμβράνης . Τμήματα της αγκύρωσης των μαστιγίων και των μαστιγίων (ο λεγόμενος δακτύλιος L) βρίσκονται επίσης στην εξωτερική μεμβράνη.

Ένα καλά μελετημένο κατά Gram-αρνητικό βακτήριο είναι η E. coli που έχει µορφή ράβδου και ανήκει στην οικογένεια των εντεροβακτηρίων (Enterobacteriaceae). Είναι ένας συµβιωτικός οργανισµός και εντοπίζεται σε τεράστιους αριθµούς στο κόλον του ανθρώπου και άλλων ζώων. Eξαιτίας του εντοπισµού του στο κόλον πολλές φορές αναφέρεται και ως κολοβακτηρίδιο. Mερικά στελέχη του βακτηρίου E.coli προκαλούν γαστρεντερίτιδα ή ουρολοιµώξεις.
https://de.wikipedia.org/wiki/%C3%84u%C3%9Fere_Membran