

Δήμητρα Σπανού , Χημικός, καθηγήτρια στο 1ο Γυμνάσιο Δάφνης
ΑΝΤΙ ΕΙΣΑΓΩΓΗΣ
Πρίν από 4 έως 3,5 δισεκατομύρια χρόνια περίπου, μέσα στις αρχέγονες θάλασσες της Γης, με διαφορετικά δεδομένα σε ότι αφορά τις συνθήκες του περιβάλλοντος, ένα "διάλυμα" από χημικές ουσίες άρχισε να ξεφεύγει από τους απλούς φυσικοχημικούς νόμους και να εκδηλώνει μια υποτυπώδη βιολογική δραστηριότητα. Ήταν ο πρώτος κρίκος στη μακριά αλυσίδα της ζωής.
Τίποτα από όσα συνέβησαν στη συνέχεια δε θα ήταν δυνατό να συμβούν, αν το αρχέγονο "διάλυμα" δεν περιβάλλοταν από ένα λεπτό περίβλημα λιποπρωτεϊνών, που το διαχώρισε από το περιβάλλον του και του επέτρεψε να διατηρήσει μια υψηλή συγκέντρωση συστατικών, κρίσιμη για την εμφάνιση της ζωής, και χωρίς αμφιβολία, έναν από τους όρους για τη διατήρησή της.
Χάρη στην πλασματική μεμβράνη, το λεπτό δηλαδή λιποπρωτεϊνικό στρώμα πάχους 8nm, τα κύτταρα αποκτούν υπόσταση διαχωριζόμενα από το περιβάλλον τους, ελέγχουν τις ανταλλαγές ουσιών με αυτό και γενικά πραγματοποιούν ένα πλήθος λειτουργιών, που θα ήταν αδύνατο να γίνουν χωρίς αυτήν.
από σχολικό βιβλίο Βιολογίας Β Λυκείου
Η ΚΥΤΤΑΡΙΚΕΣ ΜΕΜΒΡΑΝΕΣ ΤΩΝ ΠΡΩΤΩΝ ΠΡΟΚΑΡΥΩΤΙΚΩΝ ΟΡΓΑΝΙΣΜΩΝ
Η κατασκευή της στα πρώτα κύτταρα -οργανισμούς ξεκίνησε με την συνένωση λιπιδίων και την δημιουργία ενός περιτυλίγματος γύρω από τα πρώτα βιομόρια ενζύμων και RNA που είχαν αρχίσει να εξελίσσονται ήδη. Εκτός από τα λιπίδια που είναι εστέρες της γλυκερόλης, οι πρώτες κατασκευές σε αρχαιοβακτήρια 
Οι πρώτες κυτταρικές μεμβράνες αποτελούσαν απλά μια κλειστή επιφάνεια που είχε σχηματιστεί με την συμπύκνωση λιπιδίων.
Σε αρχαία συναντάμε μονοστοιβάδα που σχηματίζεται από λιπίδια και φωσφολιπίδια αιθερικών δεσμών και συμπηκνώνεται με δημιουργία τετρααιθέρες της διγλυκερόλης.
Τα λιπίδια και φωσφολιπίδια αιθερικών δεσμών δημιουργούνται με αιθεροποίηση 2 υδροξυλίων της γλυκερίνης με αλειφατικές ακλοόλες διακλαδιζόμενης αλυσίδας (π.χ. φυτανόλη) και με φωσφορικό οξύ
Μονοστοιβάδα λιπιδίων συναντάμε ακόμα και σήμερα σε κάποια θερμόφιλα βακτήρια.
Στα βακτήρια η κατασκευή των μεμβρανών γίνεται από λιπίδια που είναι τύπου εστερικού δεσμού
Η διαφορά στην κατασκευή των μεμβρανών φαίνεται ακριβώς στα διαφορετικά ποσοτά τους σε πρωτεϊνες, λιπίδια και υδατάνθρακες.
Η ΔΟΜΗ ΤΩΝ ΚΥΤΤΑΡΙΚΩΝ ΜΕΜΒΡΑΝΩΝ ΟΠΩΣ ΕΞΕΛΙΧΘΗΚΑΝ ΣΤΗΝ ΣΥΝΕΧΕΙΑ
Τα λιπίδια της κυτταρικής μεμβράνης, συμπληρώθηκαν αργότερα από πρωτεϊνες άλλες ενσωματωμένες στην μεμβράνη κι άλλες λιγότερο στενα συνδεδεμένες (περιφεριακές πρωτεϊνες).

η διπλή στοιβάδα των αμφίφιλων λιπιδίων
Η μεμβράνη των κυττάρων αποτελεί τριπλή στοιβάδα. Δύο στρώματα φωσφολιπίδιων πάχους 20Αο περίπου εξωτερικά και ενδιάμεσα ένα στρώμα από πρωτεϊνες και σάκχαρα πάχους 35Αο περίπου. Η μεμβράνη περιέχει 30% νερό και 30-50% λιπίδια και 50-70% πρωτεϊνες.
 Στα λιπίδια αυτά μια ομάδα της τρισθενούς αλκοόλης γλυκερόλης έχει εστεροποιηθεί με φωσφορικό οξύ ( φωσφολιπίδια) και παρουσιάζουν αμφίφιλο χαρακτήρα.
Στα λιπίδια αυτά μια ομάδα της τρισθενούς αλκοόλης γλυκερόλης έχει εστεροποιηθεί με φωσφορικό οξύ ( φωσφολιπίδια) και παρουσιάζουν αμφίφιλο χαρακτήρα.
 Συνήθως η ακυλομάδα του πρώτου άνθρακας της γλυκερίνης είναι κορεσμένη η ακυλομάδα του δεύτερου άνθρακα ακόρεστη ενώ η φωσφορική ομάδα είναι εστεροποιημένη με αμινοαλκοόλη όπως η χολίνη ή με αιθυλαμίνη
Συνήθως η ακυλομάδα του πρώτου άνθρακας της γλυκερίνης είναι κορεσμένη η ακυλομάδα του δεύτερου άνθρακα ακόρεστη ενώ η φωσφορική ομάδα είναι εστεροποιημένη με αμινοαλκοόλη όπως η χολίνη ή με αιθυλαμίνη
Αμφίφιλο χαρακτήρα παρουσιάζουν και άλλα λιπίδια που συμμετέχουν σε μικρότερο ποσοστό
'οπως η χοληστερόλη και τα γλυκολιπίδια
Έτσι το υδρόφιλο τμήμα ( φωσφορικής ομάδας, ΟΗ, σάκχαρα) και για τις δύο στώσεις λιπιδίων προσανατολίζεται προς τα έξω (κυτταρόπλασμα και μεσοκυκάριο χώρο) ενώ το υδρόφοβο των οργανικών αλυσίδων στο εσωτερικό της μεμβράνης. Έτσι στο εσωτερικό της μεμβράνης σχηματίζεται διπλοστοιβάδα υδρόφοβων άκρων τα οποία μπορούν να είναι κορεσμένες ή ακόρεστες. Αυτό δίνει διαφορά στην δομή τους εφόσον οι κορεσμένες είναι σφικτές και παράλληλες μεταξύ τους και κάθετες στην επιφάνεια, ενώ οι ακόρεστες είναι χαλαρές με ακανόνιστη διάταξη.
(Στα λιπίδια εντάσοσονται επίσης χοληστερόλη και γλυκολιπίδια)
Οι πρωτεϊνες της κυτταρικής μεμβράνης:
Στις μεμβράνες υπάρχουν οι κυτταρικές πρωτεϊνες που συνδέονται περιφεριακά με την κυτταρική μεμβράνη στην εξωτερική ή εσωτερική πλευρά της, πρωτεϊνες που εισέρχονται μερικώς στην μεμβράνη και πρωτεϊνες που διαπερνούν την μεμβράνη
Ως προς ρόλο τους, στην μεμβράνη συναντάμε δύο ειδών πρωτεϊνες:
τις λειτουργικές (ένζυμα, πρωτεϊνες που δρουν σαν αντλίες, μεταφορείς, πρωτεϊνες δίαυλοι (κανάλια) ιόντων) , ρυθμιστικές πρωτεϊνες και τις δομικές .
Οι λειτουργικές πρωτεϊνες "χώνονται" στο υδρόφοβο εσωτερικό της μεμβράνης και παραμένουν με υδρόφοβους δεσμούς (λιποπρωτεϊνες ) ή ακέραιες πρωτεϊνες. Δεν εκχυλίζονται.
Οι δομικές πρωτεϊνες που είναι χωρίζονται σε δύο κατηγορίες τις περιφεριακές και τις διαμεμβρανιακές.
Οι περιφεριακές πρωτεϊνες αλληλεπιδρούν με ηλεκτροστατικούς δεσμούς με τις υδρόφιλες κεφαλές των λιπιδίων ή συνδέονται με ακέραιες πρωτεϊνες με ασθενείς δεσμούς και εύκολα εκχυλίζονται χωρίς να καταστρέφεται η μεμβράνη
Υπάρχουν επίσης τέτοιες πρωτεινες στην επιφάνεια αλλά και στο εσωτερικό της μεμβράνης.
Προς την εσωτερική μεριά του κυττάρου μπορεί να συνδεθούν με ένζυμα του κυτταροπλάσματος και να πάρουν μέρος σε ενζυμικές αντιδράσεις. Ενώνονται με μικρονημάτια και μικροσωληνίσκους του κυτταροπλάσματος σε ένα πλέγμα.
Αντίστοιχα στην εξωτερική επιφάνειας του κυττάρου πρωτεϊνες και λιπίδια συνδέονται με υδατάνθρακες (γλυκοπρωτεϊνες και γλυκολιπίδια).

Οι διμεμβρανιακες πρωτεινες φροντιζουν την είσοδο και έξοδο ουσιων στο κυτταρο, σν αυτες δεν μπορούν να περάσουν αυθόρμητα δια μεσου της κυτταρικής μεμβράνης .
Οι διαμεμβρανιακές, δηλαδή αυτές που διασχίζουν την κυτταρική μεμβράνη και βγάζουν δύο υδρόφιλα άκρα, ένα στην εξωτερική και ένα στην εσωτερική πλευρά και είναι αυστηρά προσανατολισμένες και τα περισσότερα αμινοξέα τους (αμινοξικά κατάλοιπα) μέσα στο υδρόφοβο εσωτερικο της μεμβρανιακής πρέπει να είναι μη πολικά, όπως Αλανίνη, βαλίνη, Λευκίνη, Ισολευκίνη, φαιλυλαλανίνη. Τυχόν πολικές ομάδες των πεπτιδίων, (όπως CONH) συμμετέχουν σε δεσμούς υδρογόνου ώστε σχηματίζονται πτυχώσεις ή έλικες που μειώνουν την ιοντική τους επίδραση και το ενεργειακό κόστος, ώστε η διαμεμβρανιακή πρωτεϊνη να μπορεί να εισέλθει και να ενσωματωθεί στο υδρόφοβο περιβάλλον της κυτταρικής μεμβράνης
Για να γίνει αυτό πρέπει τα διαδοχικά κατάλοιπα της πολυπεπτιδικής αλυσίδας να σχηματίζουν γωνίες από -120ο έως -180ο μοίρες και από 120ο έως 180ο
Στις διαμεμβρανιακές έχουμε και η αλυσίδα αυτή δίνει μια εκτεταμένη διαμόρφωση ώστε οι αμινομάδες και καρβονυλομάδες της κύριας αλυσίδας να διατάσονται περίπου κάθετα στον άξονα της αλυσίδας για την δημιουργία δεσμών υδρογόνου με άλλα τμήματα της αλυσίδας.
Αυτές που διασχίζουν μία φορά την κυτταρική μεμβράνη και βγάζουν υδρόφιλα άκρα ένα στην εξωτερική και ένα στην εσωτερική πλευρά,
Αυτές που διασχίζουν πολλές φορές την κυτταρική μεμβράνη μπορούν να δημιουργούν διαύλους (κανάλια) για το πέρασμα ιόντων ή μορίων από τον εξωκυττάριο χώρο στο κυτταρόπλασμα και αντίστροφα (αντλία Καλίου- Νατρίου, Αντλία Ασβεστίου κ.α.)

Αντίστοιχα
α. μπορεί να διαπερνούν την λιπιδική διπλοστοιβάδα μια φορά και διαθέτουν α έλικα που βυθίζεται στον εσωτερικό χώρο που είναι η υδρόφοβη πλευρά των λιπιδίων ( π.χ. γλυκοφορίνη, μεμβρανικές ανοσοσφαιρίνες, ορμονικοί υποδοχείς, ινσουλίνη, κ.α.)
β. μπορεί να διαπερνούν την λιπιδική διπλοστοιβάδα πολλές φορές πάλι σαν α έλικα με την βοήθεια φορτισμένων αμινοξέων και παίζουν σημαντικό ρόλο στην δημιουργία διαύλων όπως η αντλία Καλίου- Νατρίου και η αντλία Ασβεστίου

Έχουμε τρείς τύπους τετοιων πρωτεϊνών: τις περιστροφικες, τις μεταφορικές και τα κσναλια. Ανεξαρτητα απο αυτό όμως,
ΣΧΗΜΑΤΙΣΜΟΣ ΤΟΥ ΓΛΥΚΟΚΑΛΥΚΑ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΟΥ ΚΥΤΤΑΡΟΥ
με την συμμετοχή γλυκοπρωτεϊνών και γλυκολιπιδίων
Οι γλυκοπρωτεϊνες και τα γλυκολιπίδια σε αυτές της επιφάνειας, συνδέονται με ίνες κολλαγόνου και δημιουργούν γύρω από το κύτταρο, μια πρόσθετη διαμόρφωση, τον γλυκοκάλυκα, που προστετεύει, μονώνει και εμποδίζει την επαφή των κυττάρων μεταξύ τους, παίρνουν μέρος στη διαβίβαση σήματος και λειτουργουν σαν αντιγόνα για το ανοσοποιητικό.
Η ΠΟΛΙΚΟΤΗΤΑ ΤΗΣ ΚΥΤΤΑΡΙΚΗΣ ΜΕΜΒΡΑΝΗΣ
Η σύσταση της εξωτερικής πλευράς της μεμβράνης προς τον εξωκυττάριο χώρο και της εσωτερικής προς το κυτταρόπλασμα διαφέρουν.
Η έξωτερικη πλευρά της μεμβράνης, έχει περισσότερα γλυκολιπίδια και χοληστερόλη λόγω της ύπαρξης του γλυκοκάλυκα, αλλά λιγότερη φωσφατιδοχολίνη κυρίως για μηχανικούς λόγους. 
Η εσωτερική πλευρά περιέχει φωσφολιπίια. Δεδομένου ότι το φωσφορικό άκρο του φωσφολιπίδιο λόγω διάστασης είναι αρνητικό το συνολικό αρνητικό φορτίο από τα φωσφολιπίδια είναι λιγότερο προς το εξωτερικό της μεμβράνης από το εσωτερικό προς το κυτταρόπλασμα και αυτό δημιουργεί ήδη μια διαφορά ηλεκτρικού δυναμικού
Εξ άλλου οι μεμβρανικές πρωτεϊνες κυτταρόπλασμα έχουν το θετικό αμινοτελικό τους άκρο στο κυτταρόπλασμα
ΠΟΙΟΙ ΟΡΓΑΝΙΣΜΟΙ ΕΧΟΥΝ ΚΥΤΤΑΡΑ ΜΕ ΠΛΑΣΜΑΤΙΚΗ ΜΕΜΒΡΑΝΗ και ΟΙ ΔΙΑΦΟΡΕΣ ΤΟΥΣ
Πλασματική εμβράνη έχουν τα κύτταρα των ιών, των φυτών, των προκαρυωτικών οργανισμών και των μυκήτων.
Γενικά οι μεμβράνες αποτελούνται από λιπίδια και πρωτεϊνες, αλλά υπάρχουν διαφορές σχετικά με το μέγεθος και τα επι μέρους συστατικά τους
Η ΔΙΑΠΕΡΑΤΟΤΗΤΑ ΤΗΣ ΚΥΤΤΑΡΙΚΗΣ ΜΕΜΒΡΑΝΗΣ
Η κυτταρική μεμβράνη είναι υπεύθυνη για τις ουσίες που εισέρχονται και εξερχονται στο κύτταρο. Υπάρχουν μόρια που εισέρχονται και εξέρχονται αυθόρμητα και άλλα που χρειάζονται την βοήθεια πρωτείνών (πρωτεϊνες μεταφορείς, πρωτείνες διαμεμβρανιακά κανάλια κ.α.).
Μεταφορά με διαβατότητα
Εφόσον στα πρώτα κύτταρα δεν υπήρχαν μεταφορικές πρωτεϊνες της μεμβράνης,
η μεταφορά ουσιών προς και από το κύτταρο γίνονταν παθητικά μέσω της μεμβράνης και όχι μέσω μεταφορικών πρωτεϊνών και με μόνο κίνητρο την κλίση των συγκεντρώσεων μέσα και έξω από το κύτταρο αφού το κύτταρο είναι βυθισμένο σε υδατώδες περιβάλλον.
Μη πολικά μόρια όπως το οξυγόνο και το διοξείδιο του άνθρακα μπορούν να διαπερνούν την κυτταρική μεμβράνη όταν υπάρχει διαφορά στις συγκεντρώσεις τους μέσα και έξω από το κύτταρο. Το νερό επίσης αν και μη πολικό μόριο διέρχεται από την κυτταρική μεμβράνη μέσω πορινών 9(ακουαπορίνες)
Μεταφορά με μεσολάβηση μεμβρανιακών πρωτεϊνων - μεταφορέων
Οι πρωτεϊνες της μεμβράνης διακρίνονται σε δομικές (που δεν απασχολεί εδώ) και λειτουργικές. Από τις λειτουργικές που είναι διαφόρων ειδών όπως ένζυμα, μεταφορείς, ειδικοί υποδοχείς.

Δήμητρα Σπανού
ΠΗΓΕΣ
ΚΥΤΤΑΡΙΚΗ ΒΙΟΛΟΓΙΑ ΛΟΥΚΑΣ ΜΑΡΓΑΡΙΤΗΣ
ΒΑΣΙΚΕΣ ΕΝΝΟΙΕΣ ΚΥΤΤΑΡΟΛΟΓΙΑΣ ΠΑΝΑΓΙΩΤΗΣ ΟΙΚΟΝΟΜΟΠΟΥΛΟΣ
https://opencourses.auth.gr/modules/document/file.php/OCRS321/%CE%A0%CE%B1%CF%81%CE%BF%CF%85%CF%83%CE%B9%CE%AC%CF%83%CE%B5%CE%B9%CF%82%20%CE%9C%CE%B1%CE%B8%CE%AE%CE%BC%CE%B1%CF%84%CE%BF%CF%82/08%2C%CE%9A%CF%85%CF%84%CF%84%CE%B1%CF%81%CE%B9%CE%BA%CE%AE%20%CE%92%CE%B9%CE%BF%CE%BB%CE%BF%CE%B3%CE%AF%CE%B18fin.pdf
Δομικά χαρακτηριστικά των μεμβρανών πλάσματος σε διάφορους οργανισμούς (thelib.info)
Клеточная мембрана Оболочка животных клеток Оболочка животной (present5.com)
ΒΙΚΙΠΑΙΔΙΑ
https://el.strephonsays.com/intrinsic-and-vs-extrinsic-proteins-13321
https://thelib.info/biologiya/984388-osobennosti-stroeniya-plazmaticheskih-membran-u-razlichnyh-organizmov/
https://dstef.weebly.com/tauiota-epsilon943nualphaiota-eta-chiomicronlambdaetasigmatauepsilonrho943nueta.html
Ακατέργαστο
αποτελεί έναν συμπαγή σχημτισμό αλλά τα μόριά που την αποτελού κινούνται το ένα προς το άλλο
Ένας βασικός ρόλος αυτών των πρωτεϊνών της μεμβράνης είναι διευκόλυνση της διαβίβασης ουσιών από έξω προς το εσωτερικό του κυττάρου
Δομές ενεργές στη μεμβράνη.
Επί του παρόντος, έχει αποδειχθεί η ύπαρξη τριών τύπων τέτοιων πρωτεϊνών μεταφοράς: κανάλια, φορείς, αντλίες.
Τα κανάλια είναι πρωτεΐνες transmembrane που δρουν σαν πόροι. Μερικές φορές ονομάζονται επιλεκτικά φίλτρα. Η μεταφορά μέσω των διώρυγας είναι συνήθως παθητική. Η ιδιαιτερότητα της μεταφερόμενης ουσίας καθορίζεται από τις ιδιότητες της επιφάνειας των πόρων. Κατά κανόνα, τα ιόντα κινούνται μέσα από τα κανάλια. Η ταχύτητα μεταφοράς εξαρτάται από το μέγεθος και τη φόρτισή τους. Εάν ο πόρος είναι ανοιχτός, τότε οι ουσίες περνούν γρήγορα. Ωστόσο, τα κανάλια δεν είναι πάντα ανοιχτά. Υπάρχει ένας μηχανισμός "πύλης" που, υπό την επήρεια εξωτερικού σήματος, ανοίγει ή κλείνει το κανάλι. Για μεγάλο χρονικό διάστημα, φαινόταν δύσκολο να εξηγηθεί η υψηλή διαπερατότητα της μεμβράνης (10 μm / s) για το νερό - μια ουσία πολική και αδιάλυτη στα λιπίδια. Επί του παρόντος, έχουν ανακαλυφθεί αναπόσπαστες πρωτεΐνες μεμβράνης, που αντιπροσωπεύουν ένα κανάλι μέσω της μεμβράνης για διείσδυση νερού - ακουαπορίνες. Η ικανότητα των ακουαπορινών να μεταφέρουν νερό ρυθμίζεται από τη διαδικασία φωσφορυλίωσης. Έχει αποδειχθεί ότι η προσκόλληση και η επιστροφή των φωσφορικών ομάδων σε ορισμένα αμινοξέα των ακουαπορινών επιταχύνει ή αναστέλλει τη διείσδυση του νερού, αλλά δεν επηρεάζει την κατεύθυνση μεταφοράς.
Οι φορείς είναι συγκεκριμένες πρωτεΐνες που μπορούν να συνδεθούν με μια ουσία που μεταφέρεται. Στη δομή αυτών των πρωτεϊνών υπάρχουν ομαδοποιήσεις με κάποιο τρόπο προσανατολισμένες στην εξωτερική ή εσωτερική επιφάνεια. Ως αποτέλεσμα των αλλαγών στη διαμόρφωση των πρωτεϊνών, η ουσία μεταφέρεται έξω ή μέσα. Δεδομένου ότι για τη μεταφορά κάθε μεμονωμένου μορίου ή ιόντων, ο φορέας πρέπει να αλλάξει τη διαμόρφωση, ο ρυθμός μεταφοράς της ουσίας είναι αρκετές φορές μικρότερος από τη μεταφορά μέσω των διαύλων. Η παρουσία πρωτεϊνών μεταφοράς όχι μόνο στο πλασμαλέμα, αλλά και στον τοννοπλάσμα παρουσιάζεται. Η μεταφορά με τη βοήθεια μεταφορέων μπορεί να είναι ενεργή και παθητική. Στην τελευταία περίπτωση, η μεταφορά αυτή πηγαίνει προς την κατεύθυνση του ηλεκτροχημικού δυναμικού και δεν απαιτεί ενεργειακό κόστος. Αυτός ο τύπος μεταφοράς ονομάζεται διευκόλυνση διάχυσης. Χάρη στους μεταφορείς, πηγαίνει με μεγαλύτερη ταχύτητα από τη συμβατική διάχυση.
ακατέργαστο
ακατεργαστο
Μεμβράνη ερυθροκυττάρων 44% , 43%, 8%,. Μυελίνη νευρώνων 18%, 79%, 3%. Ηπατοκύτταρα 54%, 36%, 10%. Πυρηνική μεμβράνη 66%, 32%, 10%. Μεμβράνες οργανιδίων Golgi 64%, 26%, 10%. Στα μιτοχόνδρια στην εσωτερική μεμβράνη 78%, 22%, 0%. Στην εξωτερική 55%, 45%, ίχνη. Μεμβράνη χλωροπλαστών 70%, 30%, 0% .

H κυτταρική μεμβράνη των ευκαρυωτικών κυττάρων όπως διαμορφώθηκε στη συνέχεια περιλαμβάνει τρεις στοιβάδες , οι δύο που έξω αποτελούνται από λιπίδια και έχουν πάχος περίπου 20 Αο ενώ η εσωτερική από πρωτεϊνες με πάχος περίπου 45 Αο.
Διαθέτει συστήματα για προσλαμβάνει και να αποβάλλει ουσίες στο κύτταρο. Βασικές τέτοιες ουσίες είναι οι στερόλες (π.χ. χοληστερόλη)
Ακατέργαστο υλικό
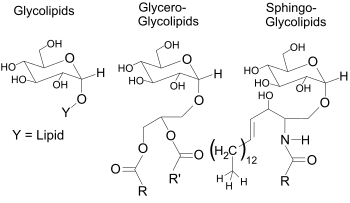
Βακτήρια: φωσφολιπίδια σε διπλή στιβάδα, εστερικοί δεσμοί, λιπαρά οξέα 16- 18 C ευθείας αλυσίδας. Αρχαία: σουλφολιπίδια, γλυκολιπίδια, μη πολικά ισοπρενοειδή, φωσφολιπίδια αιθερικοί δεσμοί, δυνατή και η μονοστιβάδα (τετραιθέρες της διγλυκερόλης σε υπερθερμόφιλα αρχαία), διακλαδιζόμενες αλκοόλες. Ο βαθμός κορεσμού των λιπιδίων εξαρτάται από τη θερμοκρασία: ψηλές θερμοκρασίες trans και κορεσμένα. Στερόλες σε μεμβράνες μερικών μυκοπλασμάτων
prokaryotes, sterols are usually absent or present in minute amounts, and they have usually been considered insignificant. However, in recent years, new classes of microbial lipids have been discovered.
πυρηνική, διπλές μεμβράνες μιτοχονδρίων και χλωροπλαστών, μεμβράνες έγκλειστων , ριβοσωμάτων, ενδοπλσματικού δικτύου.
 Τα λιπίδια εστερικών δεσμών δημιουργούνται με εστεροποίηση 2 υδροξυλίων της γλυκερίνης με καρβοξυλικά και με οξέα 16-18 ανθράκων και με φωσφορικό οξύ.
Τα λιπίδια εστερικών δεσμών δημιουργούνται με εστεροποίηση 2 υδροξυλίων της γλυκερίνης με καρβοξυλικά και με οξέα 16-18 ανθράκων και με φωσφορικό οξύ.

Η επιφάνεια της μεμβράνης δημιουργείται όταν τα λιπίδια συνδέονται μεταξύ τους με έναν βασικό χημικό μηχανισμό που ονομάζεται συμπύκνωση
Κατά τον μηχανισμό αυτόν το ένα μονομερές χάνει ένα άτομο υδρογόνου (Η), ενώ το άλλο μια υδροξυλομάδα (-ΟΗ). Δηλαδή αφαιρείται ένα μόριο νερού.
Έτσι, οι πρώτες κυτταρικές μεμβράνες αποτελούσαν απλά μια κλειστή επιφάνεια που είχε σχηματιστεί με την συμπύκνωση λιπιδίων.
Σε αρχαία συναντάμε μονοστοιβάδα που σχηματίζεται από τετρααιθέρες της γλυκερόλης Μονοστοιβάδα λιπιδίων συναντάμε ακόμα σε κάποια θερμόφιλα βακτήρια,
 ενώ, σε επόμενη φάση σε αρχαία και βακτήρια η κυτταρική μεμβράνη σχηματίζεται με διπλοστοιβάδα λιπιδίων
ενώ, σε επόμενη φάση σε αρχαία και βακτήρια η κυτταρική μεμβράνη σχηματίζεται με διπλοστοιβάδα λιπιδίων
Τα λιπίδια αυτά στην διπλοστοιβάδα, είναι εστερικής δομής και με την φωσφορική ομάδα με ένα άλλο μικρό μόριο να είναι το υδρόφιλο μέρος καιπρος τα έξω που έρχεται σε επαφή με μόρια νερού του πλάσματος και του εξωτερικού περιβάλλοντος, ενώ
H συνένωση των λιπιδίων αυτών γίνεται με συμπύκνωση και όχι πολυμερισμό και η εικόνα που δίνει η μεμβράνη λιπιδίων από την έξω πλευρά είναι σωματιδιακού χαρακτήρα ενώ στην άλλη πλευρά οι αλυσίδες των λιπαρών οξέων που εστεροποιούνται δίνουν την εικόνα ενός πλέγματος από ινίδια.
.png)
Η χημική σύσταση των λιπιδίων αυτών στα μεν αρχαία ήταν σουλφολιπίδια, γλυκολιπίδια, μη πολικά ισοπρενοειδή,
Στα βακτήρια τα λιπίδια ήταν φωσφολιπίδια και λιπαρά οξέα με αριθμό ανθράκων 16 έως 18 , αριθμός που είναι σχετικός με τα λιπαρά οξέα που συμμετείχαν σε πιο εξελιγμένα κύτταρα στην συνέχεια
Τα λιπαρά οξέα που εστερποιούνται σε λιπίδια της μεμβράνης, μπορεί να είναι κορεσμένα ή ακόρεστα πράγμα που εξαρτάται από την θερμοκρασία. Σε υψηλές θερμοκρασίες (ηφαίστεια, ραδιενεργά κοιτάσματα) όπου επικρατούν οξειδωτικές συνθήκες είναι κορεσμένα (δηλαδή χαμηλότερης ενέργειας). Σε κορεσμένα λιπίδια οι αλυσίδες των λιπαρών οξέων συνδέονται σφικτά και παράλληλα μεταξύ τους και κάθετα στην επιφάνεις. Συνολικά δίνουν ένα κύτταρο με σκληρή κυτταρική μεμβράνη
Αντίθετα σε μεταγενέστερα βακτήρια της επιφάνειας που επικρατούσε χαμηλώτερη θερμοκρασία και αναγωγικές συνθήκες, πιθανόν να ![]() έχουμε και ακόρεστα σε διατάξεις trans αλλά και cis στις οποίες δημιουργούν λιπίδια με χαλαρή διάταξη και συνολικά πιο εύκαμπτη μεμβράνη
έχουμε και ακόρεστα σε διατάξεις trans αλλά και cis στις οποίες δημιουργούν λιπίδια με χαλαρή διάταξη και συνολικά πιο εύκαμπτη μεμβράνη
Τα πρώτα προκαρυωτικά κύτταρα δεν έχουν στερόλες στις κυτταρικές τους μεμβράνες, -οι οποίες είναι μεταξύ άλλων επιφορτισμένες να εισάγουν επιλεκτικά ουσίες στο κύτταρα. Η είσοδος και έξοδος ουσιών από το κύτταρο γίνεται παθητικά με διάχυση
Όμως σε Έχουν διπλή σειρά λιπιδίων (διπλοστοιβάδα).
(Αυτή διατηρήθηκε και στην συνέχεια της εξέλιξης των ειδών αλλά εκεί προσθέθηκαν επιπλέον και οι κυτταρικές πρωτείνες).
.jpg)

ακατεργαστο