
Ττης Δήμητρας Σπανού, Χημικού, μόνιμης καθηγήτριας Δευτ/θμιας Εκπ/σης 1ου Γυμν. Δάφνης
υπό κατασκευή
ΒΙΟΣΥΝΘΕΣΕΙΣ ΑΠΟ ΜΕΤΑΒΟΛΙΤΕΣ ΤΟΥ ΚΥΚΛΟΥ ΚΡΕΜΠΣ
.JPG)
Ο κύκλος του Κρεμπς εκτός από τον βασικό μεταβολικο κύκλο του καταβολισμού λειτουργεί και σαν δίσκος περιστροφής . Από τους μεταβολίτες του Κυκλου Κρεμπσ περνάμε σε αναβολικές πορείς από τις οποίες βιοσυντίθενται και εξασφαλίζονται οι απαραίτητες για τον οργανισμό οργανικές ουσίες.
Βιοσυνθέσεις από το ακέτυλοσυνένζυμο Α
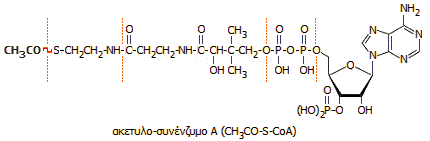
Εδώ θα δούμε τα προιόντα που παίρνουμε στην αρχή του κύκλου από το ακέτυλοσυνένζυμοΑ.
Το ακέτυλο συνένζυμο Α προέρχεται από την γλυκόλυση αλλά και την διάσπαση λιπαρών οξέων .
Το οξύ που συνδέεται με το συνένζυμοΑ είναι το οξικό οξύ και γι αυτό μπορούμε να το θεωρήσουμε σαν το ενεργοποιημένο οξικό οξύ.
Πλην των πολλών βιοσυνθέσεων όπως των λιπαρών οξέων και των στερολών το συναντούμε και σε άλλες περιπτώσεις εφόσον, είναι ο βασικός φορέας και δωρητής ακυλοομάδων στο σώμα,
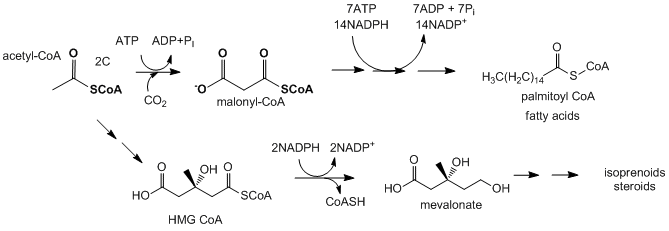
Για τις συνθέσεις από το ακέτυλο CoA χρειάζεται ενέργεια (ΑTP) και ανάλογα ένζυμα.
Η βιοσύνθεση λιπαρών οξέων στους ζωικούς οργανισμούς
Η βιοσύνθεση λιπαρών οξέων στους ζωικούς οργανισμούς γίνεται στο κυτταρόπλασμα.
Όμως το ακέτυλοσυνένζυμοΑ παράγεται στα μιτοχόνδρια και για να χρησιμοποιηθεί για την παραγωγή λιπαρών οξέων πρέπει να εξέλθει από τα μιτοχόνδρια πράγμα που θα το πετύχει μέσω του κιτρικού οξέος
Το κιτρικό οξύ εγκαταλείπει τα μιτοχόνδρια , περνά στο κυτταρόπλασμα και διασπάται πάλι σε ακέτυλοσυνένζυμο και οξαλοξικό με το ένζυμο ATPκιτρική λυάση. Η ενέργεια που χρειάζεται την παίρνει από την διάσπαση ενός μορίου ATP.
Το οξαλοξικό ανάγεται πάλι σε μηλικό και διαπερνά αντίθετα την μιτοχονδριακή μεμβράνη για να συμμετέχει στον κύκλο Κρεμπς.
Εάν υπάρχει ένδεια σε ΑΤP ο κύκλος του κιτρικού οξέος προχωρά ταχέως και δεν εξέρχεται κιτρικό οξύ από το κυτταρόπλασμα που θα ξεκινήσει την παραγωγή λιπαρών οξέων. Εάν όμως υπάρξει περίσσεια ΑΤP ο κύκλος του κιτρικού επιβραδύνεται και εξέρχεται κιτρικό από το μιτοχόνδριο προς το κυτταρόπλασμα για την βιοσύνθεση των λιπαρών οξέων.
Η προετοιμασία της σύνθεσης των λιπαρών οξέων
Σύνδεση με το CoA. Ενεργοποίηση του οξικού και μηλονικού οξέος.
 H Αντίδραση σχηματισμού κιτρικού ελευθερώνει ενέργεια -32,2kj/mol. Επομένως η αντίδραση διάσπασης του κιτρικού που προέρχεται από τον κύκλο του Κρεμπς είναι 32,2kj/mol και προέρχεται
H Αντίδραση σχηματισμού κιτρικού ελευθερώνει ενέργεια -32,2kj/mol. Επομένως η αντίδραση διάσπασης του κιτρικού που προέρχεται από τον κύκλο του Κρεμπς είναι 32,2kj/mol και προέρχεται
από την διάσπαση ενός μορίου ATP
Σχηματισμός μηλονικού οξέος : Στην βιοσύνθεση λιπαρών οξέων στο ακέτυλοσυνένζυμοΑ που βρίσκεται στο κυτταρόπλασμα, προστίθεται αρχικά CO2 με ένζυμο την βιοτίνη
και σχηματίζεται μηλονικο οξύ συνένζυμοΑ (μηλονιλοσυνένζυμοΑ). Το μόριο αυτό αντιδρά πιο έντονα και πιο ευνοικά για την επιμήκυνση της αλυσίδας. Η αντίδραση γίνεται με κατανάλωση ενός μορίου ΑΤΡ
![]()
Η αρχή της σύνθεσης των λιπαρών οξέων
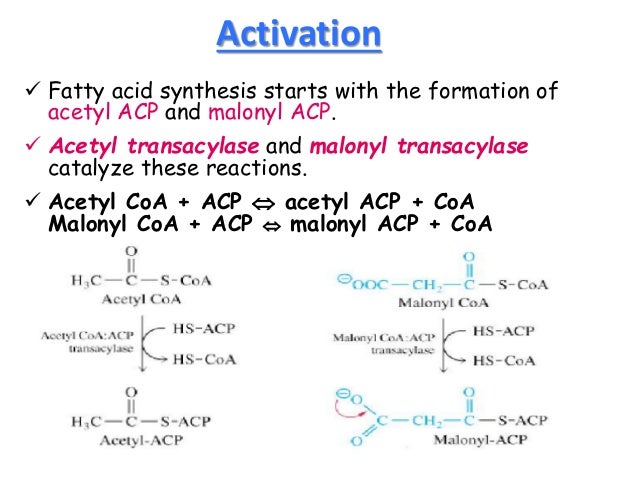
Στην αρχή της σύνθεσης αφού ενεργοποιηθούν τα οξέα με την ένωσή τους με το CoA , σε ένα μόριο ακέτυλοσυνένζυμοΑ και ένα μαλονυλσυνένζυμοΑ ενώνονται μεταξύ τους στην συνέχεια προς ακετοακέτυλοCoA .
Το ένζυμο και ο δότης διττανθρακικών μονάδων για σχηματισμό- επιμήκυνση αλυσίδας
Στον σχηματισμό των λιπαρών οξέων συμμετέχει ένα πολυενζυμικό σύμπλεγμα που λέγαται συνθάση των λιπαρών οξέων.
Για τις αντιδράσεις που καταλύει η συνθάση, όλα τα ενδιάμεσα προιόντα έρχονται συνδεμένα με μια μικρή πρωτείνη την Ακετυλ - φέρουσα πρωτείνη (ACP-SH) που είναι κομμάτι της συνθάσης των λιπαρών οξέων. Η ένωση δύο ουσιών για την σύνθεση γίνεται στην στη -SH της ACP
Το μηλονιλοσυνένζυμοΑ είναι ο σταθερός δότης για την προσθήκη μονάδων από δυο άνθρακες ώστε να σχηματιστεί σταδιακά η αλυσίδα των λιπαρών κορσμένων οξέων
Η επιμήκυνση της αλυσίδας των λιαρών οξέων
Οι ενώσεις που χρειάζονται, για να συναντηθούν, μεταφέρονται κοντά με την βοήθεια των ενζύμων Ακετυλ - CoA ACP - transacetylase και Μαλονυλ - CoA transferase αφού ενώνονται πρώτα με την ACP στην συνθάση των λιπαρών οξέων
Το αποτέλεσμα είναι ο σχηματισμός μιας μονάδας ακυλίου τεσσάρων ατόμων άνθρακα ενωμένου με CoA το ακεοακέτυλο CοΑ που αν υδρολυθεί θα προκύψει το οξα παλμιτικό οξύ
 Η σύνθεση του παλμιτικού CoA γίνεται σταδιακά σε τέσσερα βήματα
Η σύνθεση του παλμιτικού CoA γίνεται σταδιακά σε τέσσερα βήματα
1. Ένωση C-C με απλό δεσμό
2.Αναγωγή
3. Αφυδάτωση
4. Μια αναγωγική αντίδραση
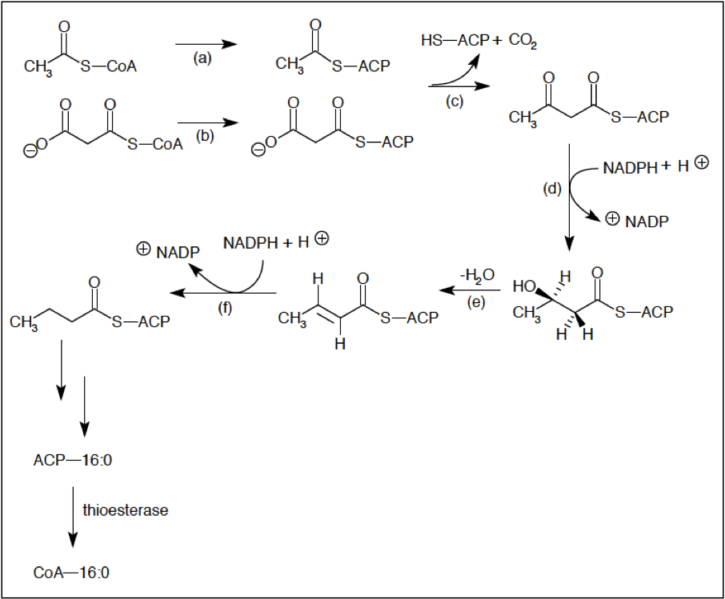
Η διαδικασία επαναλαμβάνεται με την μεταφορά ενός νέου ενεργοποιημένου μορίου οξικού στο ακέτυλοσυένζυμοΑ
Oi ελεύθερες ενέργειες για τις αντιδράσεις του σχηματισμού λιπαρών οξέων δίνονται κάτω:
ATP + H2O --> ADP + ανόργανο φωσφορικό ΔG= -30,5 kj/mol (Ph=7, Θερμοκρασία 25οC)
ATP + H2O -> AMP + PPi ΔG°ʹ = –45,6 kJ mol–1 (–10,9 kcal mol–1)
Ακετυλο-CoA + Η2Ο -> οξικό + CoA + Η+ ΔG°ʹ = –31,4 kJ mol–1 (–7,5 kcal mol–1)
ΣΧΗΜΑΤΙΣΜΟΣ ΤΡΙΓΛΥΚΕΡΙΔΙΩΝ
Δημιουργούνται με την ένωση των λιπαρών οξέων με την τρισθενή ακλοόλη γλυκερίνη
Σε πρώτο στάδιο η γλυκερίνη ενεγοποιείται με την φωσφορυλίωσή της από το ATP και με κατανάλωση ενός μορίου ATP.

Στην συνέχεια το αγλυκερινοφωσφορικό οξύ αντιδρά με λιπαρά οξέα ενεργοποιημένα με ακέτυλοσυνένζυμοΑ και
με υδρόλυση ενός μορίου ATP σε AMP

Στην συνέχεια το αγλυκερινοφωσφορυλικό οξύ αντιδρά με λιπαρά οξέα ενεργοποιημένα με CoA και εστεροποιούνται έτσι ο 3ος και στην συνέχεια με ένα δεύτερο μόριο οξέος ο δεύτερος άνθρακας του μορίου
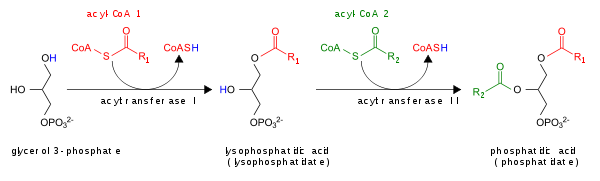
'Ετσι σχηματίζεται το φωσφατιδικό οξυ που είναι πρόδρομη ένωση πολλών λιπιδίων και των τριγλυκεριδίων

Oi ελεύθερες ενέργειες για τις αντιδράσεις του σχηματισμού τριγλυκεριδίων δίνονται κάτω:
ATP + H2O --> ADP + ανόργανο φωσφορικό ΔG= -30,5 kj/mol (Ph=7, Θερμοκρασία 25οC)
ATP + H2O -> AMP + PPi ΔG°ʹ = –45,6 kJ mol–1 (–10,9 kcal mol–1)
Ακετυλο-CoA + Η2Ο -> οξικό + CoA + Η+ ΔG°ʹ = –31,4 kJ mol–1 (–7,5 kcal mol–1)
ΠΗΓΕΣ
https://www.biology.uoc.gr/courses/BIO392_efarmbio/2liparwn.htm
https://el.wikipedia.org/wiki/%CE%9B%CE%B9%CF%80%CE%B1%CF%81%CF%8C_%CE%BF%CE%BE%CF%8D
https://en.wikipedia.org/wiki/Fatty_acid_synthesis#/media/File:Acety-CoA_ACP_transacylase.png
https://employees.csbsju.edu/hjakubowski/classes/ch331/Metabolism/MetPathways2_Anabolilsm.html
Σύνθεση των λιπαρών οξέων στον ’νθρωπο
Γίνεται στο κυτταρόπλασμα. Η σύνθεση των λιπαρών οξέων αρχίζει από το Ακετυλ - CoA το οποίο προέρχεται από τη γλυκόλυση ή από την αποικοδόμηση των λιπαρών οξέων· και στις δύο περιπτώσεις σχηματίζεται στα μιτοχόνδρια και χρειάζονται ειδικοί μεταβολικοί δρόμοι για την έξοδό τους απ' αυτά. Το τελικό προϊόν του ενζύμου Λιπαρό οξύ - συνθάση είναι το παλμιτικό οξύ. Στο παλμιτικό οξύ μόνο τα άτομα άνθρακα 15 και 16 προέρχονται από Ακετυλ - CoA ενώ τα υπόλοιπα προκύπτουν από το Μαλονύλ - CoA. Το Μαλονυλ - CoA προέρχεται επίσης από το Ακετυλ - CoA με τη δράση του ενζύμου Ακετυλ - CoA Καρβοξυλάση. Το ένζυμο αυτό έχει σαν συνένζυμο τη βιοτίνη.
![]()
Το ένζυμο Ακετυλ - CoA Καρβοξυλάση καθορίζει την ταχύτητα της σύνθεσης του παλμιτικού οξέος και αναστέλλεται από το τελικό προϊόν.
Όλα τα ενδιάμεσα προϊόντα της σύνθεσης των λιπαρών οξέων είναι συνδεμένα σε μια μικρή πρωτείνη που ονομάζεται Ακετυλ - φέρουσα πρωτείνη (ACP-SH) και είναι ένα κομμάτι της Συνθάσης των λιπαρών οξέων. Η Ακετυλ - φέρουσα πρωτείνη έχει δηλαδή ιδιότητες ανάλογες του CoASH. Τα ενδιάμεσα προϊόντα της σύνθεσης του λιπαρού οξέος είναι συνδεμένα με το SH της πρωτείνης υπό μορφή θειοεστέρα.
Για την έναρξη της σύνθεσης των λιπαρών οξέων ένα Ακετυλ - CoA και ένα Μαλονυλ - CoA πρέπει πρώτα να ενωθούν με την ACP στη συνθάση των λιπαρών οξέων: Η ένωση των δύο αυτών ουσιών γίνεται στη -SH της ACP του ενζύμου της συνθάσης των λιπαρών οξέων. Η ένωση του Ακετυλ - και του Μαλονυλ - στη συνθάση επιτυγχάνεται με τα ένζυμα Ακετυλ - CoA ACP - transacetylase και Μαλονυλ - CoA transferase. Έτσι οι δυο πρόδρομες ουσίες για τη σύνθεση των λιπαρών οξέων είναι ενεργοποιημένες και είναι έτοιμες για την ένωσή τους για το σχηματισμό της πρώτης ουσίας με 4 άτομα άνθρακα. Αυτό επιτυγχάνεται με ένα κύκλο από τέσσερις αντιδράσεις. Η πρώτη αντίδραση είναι ένωση άνθρακα - άνθρακα με απλό δεσμό. Η δεύτερη είναι μια αναγωγή, η τρίτη μια αφυδάτωση και η τέταρτη είναι μια αναγωγική αντίδραση. Τα ένζυμα που είναι υπεύθυνα είναι το β - Κετοακυλ - ACP συνθάση, η β - Κετοακυλ - ACP ρεδουκτάση, η 3 - Υδροξυακυλ - ACP αφυδατάση και η Ενουλ ACP ρεδουκτάση. Έτσι για τη σύνθεση του Παλμυτουλ ACP γίνονται 7 τέτοιοι κύκλοι. Μετά με υδρόλυση απελευθερώνεται το παλμιτικό οξύ από την ACP. To υδρολυτικό ένζυμο παλμιτουλ - θειοεστεράση είναι ένα μέρος του ενζύμου της συνθάσης του Λιπαρού οξέος.
Λιπαρά οξέα που έχουν περισσότερα των 16 ατόμων C συντίθενται στο ενδοπλασματικό δίκτυο. Η αντίδραση αυτή είναι όμοια με τις προηγούμενες με νέα προσθήκη δύο ατόμων C σε τύπο του Μαλονυλ - CoA. Όλα όμως τα ενδιάμεσα προϊόντα ενεργοποιούνται με το CoASH και όχι με ACP.
Βιοσύνθεση των ακόρεστων λιπαρών οξέων
Τα ακόρεστα λιπαρά οξέα συντίθενται στο ενδοπλασματικό δίκτυο με τα ένζυμα που λέγονται Ακυλ - CoA δεσατουράσες. Ένα παράδειγμα αποτελεί η σύνθεση του ελαιοΰλ - CoA από το στεαρυλ - CoA.
![]()
Τα δύο επόμενα λιπαρά οξέα το λινολεικό (18:2 Δ9, 12) και το λινολενικό (18:3 Δ9, 12, 15) είναι σημαντικά για την ανάπτυξη του ανθρώπινου οργανισμού δεν μπορούν να συντεθούν από τον ανθρώπινο οργανισμό από τις τροφές. Το λινολενικό είναι απαραίτητο συστατικό των σφυγγολιπιδίων του δέρματος και πρόδρομος ουσία στο σχηματισμό του αραχιδονικού. Το αραχιδονικό είναι πρόδρομος ουσία της προσταγλαδίνης και θρομβοξάνης.
Το λινολεϊκό οξύ είναι απαραίτητο συστατικό της τροφής του ανθρώπου. Η εκτίμηση που υπάρχει σήμερα είναι ότι ένα κανονικό άτομο χρειάζεται περίπου 10g/ημέρα λινολενικό οξύ. Το λινολεϊκό οξύ έχει δύο διπλούς δεσμούς στη θέση Δ9 και Δ12. Από το λινολεϊκό οξύ προκύπτουν δύο παράγωγα με τρεις διπλούς δεσμούς. Το ένα έχει τον ένα επιπλέον δεσμό στη μετά το Δ12 θέση δηλαδή στη Δ15. Αυτό το προϊόν λεγεται α-λινολενικό οξύ και βρίσκεται σε όλα σχεδόν τα φυτά και το άλλο έχει το διπλό δεσμό προς την καρβοξυλική ομάδα και δηλαδή στη θέση Δ6 και λέγεται γ-λινολενικό οξύ. Αυτό το γ-λινολενικό οξύ (Δ6, 9, 12) απαντά σε ελάχιστα ποσά στους ανθρώπινους και ζωικούς ιστούς και αυτό θεωρείται πρόδρομη ουσία για το Αραχιδονικό οξύ. Το αραχιδονικό οξύ έχει C20 και φέρει 4 διπλούς δεσμούς προέρχεται από το γ-λινολενικό οξύ με μια επιμήκυνση που ακολουθείται από μια desaturation για το σχηματισμό του διπλού δεσμού στη θέση Δ5. το αραχιδονικό έχει 4 διπλούς δεσμούς C20(Δ5, 8, 11, 14
Σύνθεση λιπαρών οξέων στα φυτά
Η βιοσύνθεση των αποταμιευτικών λιπιδίων στους καρπούς των φυτών συμπεριλαμβάνει τη συνεργασία των Λευκοπλαστών (πλαστίδια των αναπτυσσόμενων σπόρων), του κυτοπλάσματος και του ενδοπλασματικού δικτύου. Τα Μιτοχόνδρια εφοδιάζουν την πρόδρομη ουσία το Ακετικό. Οι Λευκοπλάστες είναι τα οργανίδια όπου η σακχαρόζη μετατρέπεται σε Ακετυλ - CoA. Το Ακετυλ - CoA είναι η πρωταρχική ουσία για τη σύνθεση των λιπαρών οξέων με την παροχή των δύο πρώτων ατόμων άνθρακα (από την η-σειρά). Ο μηχανισμός του σχηματισμού των λιπαρών οξέων στα πλαστίδια διαφέρει από ιστό σε ιστό και από το στάδιο ανάπτυξης του ιστού. Γενικά υπάρχουν δύο βιοσυνθετικοί δρόμοι: 1) η κυτοπλασματική Σακχαρόζη διασπάται σε εξόζη - φωσφορική και μετά εισέρχεται στους Λευκοπλάστες και εκεί μετατρέπεται σε Ακετυλ - CoA δια μέσου της γλυκόλυσης που γίνεται στους Λευκοπλάστες. 2) η κυτοπλασματική σακχαρόζη διασπάται σε πυροσταφυλικό οξύ (γλυκόλυση) και μετά εισέρχεται στους Λευκοπλάστες και μετατρέπεται σε Ακετυλ - CoA. Ελεύθερο ακετικό οξύ εισερχόμενα στους λευκοπλάστες από τα μιτοχόνδρια μετατρέπονται σε Ακετυλ - CoA με το ένζυμο Ακετυλ - CoA των πλαστιδίων. Ο μηχανισμός που μεταφέρει το Ακετικό οξύ από τα Μιτοχόνδρια είναι το σύστημα της L - ακετυλκαρνιτίνης / καρνιτίνη.
Η βιοσύνθεση των λιπαρών οξέων αρχίζει με τη σύνθεση του Μαλονυλ - CoA από το Ακετυλ - CoA με το ένζυμο Ακετυλ - CoA Καρβοξυλάση.
Μετά από αυτό αρχίζουν μια σειρά συνδετικών αντιδράσεων που καταλύονται με το σύμπλοκο ένζυμο τη συνθετάση των λιπαρών οξέων (FAS). Το σύμπλοκο αυτό περιλαμβάνει τις β-κετοακυλ-ACP συνθάσες (KASs) που δημιουργούν τη σύνθεση των κορεσμένων λιπαρών οξέων στους λευκοπλάστες. Το τελικό λιπαρό οξύ που συντίθεται στα φυτά είναι το στεαρικό οξύ (18:0).
Μια ακυλ-φέρουσα πρωτείνη (ACP) είναι το συνένζυμο που συνδέεται με την αύξηση της υδρογονανθρακικής αλυσίδας του λιπαρού οξέος. Στεαρουλ-ACP μετατρέπεται μετά σε ελαιοΰλ - ACP με τη διαλυτή δεσατουράση των λευκοπλαστών τη Δ9 - στεορουλ-ακυλ φέρουσα πρωτείνη δεσατουράση.
Μια θειοεστεράση η Ελαιοΰλ - ACP θειοστεράση των πλαστιδίων χωρίζει το ACP μόριο και ελευθερώνεται το ελαιοΰλ - CoA. Αυτό είναι το τελικό προϊόν της σύνθεσης στους λευκοπλάστες που αυτό δημιουργεί το Ακυλ - CoA που αποταμιεύεται στο ενδοπλασματικό δίκτυο για να συνθέσει τα τριγλυκερίδια.
Το Ελαιοΰλ - CoA είναι επίσης το υπόστρωμα για διάφορα συνθετικά συστήματα που δημιουργούν τη σύνθεση των λιπαρών οξέων με μεγαλύτερα μόρια στο κυτταρόπλασμα. Το ελαιοΰλ - CoA μεταφέρεται στο ενδοπλασματικό δίκτυο πιθανόν με μια ομάδα πολυπεπτιδίων που ονομάζονται λιπίδια μεταφορικές πρωτείνες (LTPs).
H σουκρόζη είναι επίσης το υπόστρωμα της σύνθεσης της γλυκερόλ - 3 -φωσφορικής που είναι η ουσία που συνθέτει τα τριγλυκερίδια. Η γλυκερόλ - 3 - φωσφορική συντίθεται στο κυτταρόπλασμα και μεταφέρεται μετά στο ενδοπλασματικό δίκτυο. Τα τριγλυκερίδια μετά μεταφέρονται στα ελαιοστόματα των σπορίων και εκεί συσσωρεύονται.
Ακελυλ - CoA Καρβοξυλάση (ACCase)
Είναι το ένζυμο που μετατρέπει το Ακετυλ - CoA σε Μαλονυλ - CoA κατά την αντίδραση:
Είναι ουσιαστικά υπεύθυνο για το σχηματισμό όλου του ανθρακικού σκελετού του λιπαρού οξέος. Όπως είναι γνωστό σε κάθε κύκλο προστίθενται δύο άτομα άνθρακα στην αναπτυσσόμενη αλυσίδα από το Μαλονυλ - CoA.
Το ένζυμο αυτό υπάρχει σε πολυενζυματική και πολυλειτουργική μορφή σε όλα τα μονο - και δι- κοτυλήδονα.
Έχει ρυθμιστική δράση. Παράγοντες που το ρυθμίζουν είναι το φως, καθώς και το στάδιο ανάπτυξης του καρπού. Είναι περισσότερο ενεργό με το φως και η ενεργότητα του προηγείται της απόθεσης των λιποειδών στον καρπό ή το σπέρμα. Αυτό το κάνει να διαφέρει από τα υπόλοιπα ένζυμα της σύνθεσης των λιπαρών οξέων που είναι ενεργά κατά τη διάρκεια της απόθεσης των λιποειδών. Καρποί που παρουσιάζουν μεγάλη απόθεση λιποειδών έχουν περισσότερο ενεργό ένζυμο από εκείνα που αποταμιεύουν μικρά ποσά λιποειδών. Σαν πολυένζυμο αποτελείται από τα ετερομερικά πεπτίδια BC, BCCP, TCa και TCβ, που κωδικοποιούνται από διαφορετικά γονίδια. Αυτό το κάνει να παρουσιάζει αρκετές δυσκολίες σε περίπτωση που θέλουμε να κάνουμε τρανσγενικά φυτά.
Μαλονυλ - CoA - ACP Τρανσακυλάση (M CAT)
Το Μαλονυλ - CoA-ACP-Tρανσακυλάση είναι το ένζυμο που καταλύει τη σύνδεση του Μαλονυλ - CoA με την ACP. Το ένζυμο αυτό βρίσκεται σε αφθονία σε όλους τους οργανισμούς και δεν απαιτείται επέμβαση με μοριακές τεχνικές. Αυτά έχουν δείξει πειράματα με την απομόνωση του γονιδίου του ενζύμου από την Ε. Coli (fabD gene) και τη μεταφορά του σε ντομάτα ή καπνό.
Η σύνθεση των λιπαρών οξέων δεν μεταβάλλεται

Ακετυλ - CoA - ACP Τρανσακυλάση (ACAT)
Το ένζυμο (ACAT) καταλύει την αντίδραση της σύνδεσης του Ακετυλ - CoA με τη ACP. Το ένζυμο αυτό δεν έχει καμία ρυθμιστική ικανότητα όπως δείχνουν πειράματα που έγιναν σε απομονωμένους χλωροπλάστες που επωάσθηκαν με ραδιενεργό 14C

Β- Κετοακυλ - ACP Συνθάση (KASs)
Τρεις ισοforms του ενζύμου υπάρχουν στα φυτά για τη σύνθεση των λιπαρών οξέων (KASI, KASII, KASIII).Η σύνδεση του Ακετυλ - CoA με το μαλονυλ - ACP για το σχηματισμό 4 ατόμων άνθρακα με τον isoform ΚΑSIII. O ισοform KASI θεωρείται ότι συμμετέχει για την παραγωγή των C6 μέχρι C16. Ο τρίτος τύπος ο KASII το σχηματισμό του Stearyl - ACP από το palmitoyl - ACP. Ο τύπος KASI αναστέλλεται με τη cerulinin σε μικρή δόση ενώ KASII είναι λιτότερο ευαίσθητος. Ο KASIII δεν αναστέλλεται καθόλου από τη cerulinin.
To ένζυμο KASIII έχει βρεθεί στην Ε. Coli. Υπερέκφραση αυτού του γονιδίου ελαττώνει την παραγωγή του 18:1 και αυξάνει την παραγωγή του μικρότερου σε αριθμό ανθράκων λιπαρών οξέων όπως του μυριστικού οξέος (14:0). Τη δράση του ενζύμου αυτού παρατηρούμε και όταν το γονίδιο που είναι υπεύθυνο για τη σύνθεση του ένζυμου εκφράσθηκε στη Brasica napus.
Κορεσμένα λιπαρά οξέα
Μια αλλαγή στο ποσό των κορεσμένων λιπαρών ή στο μήκος της ανθρακικής αλυσίδας έχει εμπορική αξία. Τα λιπαρά οξέα C20 - C24 έχουν εμπορική αξία σαν λιπαντικά, διαλυτικά και συνθετικά.
Τα λιπαρά οξέα C6-C14 χρησιμοποιούνται στη σαπωνοποιία σε απορρυπαντικά και πλαστικά.
Τα λιπαρά οξέα C16-C18 χρησιμοποιούνται στη μαργαρίνη, στα βρώσιμα γενικώς παρασκευάσματα από ψωμιά και γάλα.
Τα φυτικά έλαια αντικαθιστούν τα ζωικά λίπη στην ανθρώπινη διατροφή. Τα φυτικά έλαια δίδουν ένα 15-20% των συνολικών θερμίδων που παίρνει ο άνθρωπος. Το ήμισυ της συνολικής χρήσης των φυτικών ελαίων καταναλώνονται σε μορφή μαργαρίνης όπου τα κορεσμένα λιπαρά οξέα αποτελούν την πρωταρχική ύλη. Σήμερα τα κορεσμένα λιπαρά οξέα παρασκευάζονται χημικά με υδρογονάση. Μερική υδρογόνωση μεταβάλλει τη σχέση κορεσμένα: ακόρεστα με αποτέλεσμα να μεταβάλλονται τα φυσικοχημικά χαρακτηριστικά των ελαίων.
Από την άλλη μεριά τα trans - λιπαρά οξέα που σχηματίζονται κατά τη διαδικασία της υδρογόνωσης. Τα trans - λιπαρά οξέα αν και σε ορισμένα στάδια της ανάπτυξης του ανθρώπου έχουν μια βιολογική σημασία είναι γενικώς επικίνδυνα γιατί αυξάνουν τη χοληστερόλη, μεταβάλλουν τη λιποπρωτείνη του αίματος βοηθούν τη θρόμβωση και είναι συνδεδεμένα με καρδιακές παθήσεις.
Η αντικατάσταση επομένως των υδρογονωμένων λιπαρών οξέων με φυσικά κορεσμένα ελαττώνει την πιθανότητα πρόσληψης trans - λιπαρά οξέα. Αυτό μπορεί να επιτευχθεί με τρανσγενικά φυτά τα οποία μπορούν να παράγουν κορεσμένα λιπαρά οξέα. Το 1992 παράχθηκαν με αυτό τον τρόπο φυτά της Brasica που παράγουν κορεσμένα λιπαρά οξέα με τη μέθοδο του μπλοκαρίσματος του ενζύμου στεαρυλ ACP δεσατουράση (SAD).
Μια άλλη έρευνα έδειξε ότι με υπερέκφραση μιας θειοεστεράσης από το αυξάνει το στεαρικό οξύ κατά 11%. Η έκφραση και των δύο ενζύμων μαζί μπορεί να αυξήσει το στεαρικό οξύ κατά 45%.
Το Λαουρικό οξύ (C12:0) είναι ιδεώδες για το σχηματισμό επιφανειών και τούτο γιατί διαλύεται τόσο στο νερό όσο και οργανικούς διαλύτες. Η κύρια πηγή του Λαουρικού οξέος είναι φοινικέλαιο και το κοκοέλαιο που αναπτύσσονται κάτω από τροπικές συνθήκες. Το λαουρικό οξύ παράγεται σήμερα από τρανσγενικά φυτά. Τούτο επετεύχθη με απομόνωση του γονιδίου από Umbellularia colifornica στο οποίο το μεγαλύτερο ποσοστό των λιπιδίων είναι το C12:0 και μεταφυτεύθηκε στο Αrabidopsis και το canola (είδη Brasica). To ποσό του C12:0 αυξήθηκε στο 25% στο Arabidopsis και στο 45% στο Kanoli. Τα φυτά αυτά ήταν εξίσου αποδοτικά όπως τα κανονικά. Ανάλογα πειράματα γίνονται για την παραγωγή και άλλων μεσαίων λιπαρών οξέων όπως του μυριστικού και καπρικού κ.α.